DECODING THE TRACE OF LIFE IN THE PRECAMBRIAN STRATA
Shin-ichi Kawakami1 and Bunji Tojo2
1 Faculty of Education, Gifu University, Gifu, 501-1193, Japan
2 Graduate School of Environmental Studies, Nagoya University, Nagoya, 464-8602, Japan
(Received 15 December 2004 Accepted 17 December 2004)
(Abstract)
Major events concerning the early evolution of life have occurred in the pre-Cambrian era. Quality and quantity of the fossil records with important information regarding the early evolution of life have been insufficient. The introduction of geochemical methods and the recent discoveries of well-preserved fossils that have provided high resolution data for the discussions on the early evolution of life are reviewed. The major events of the early evolution of life are the emergence of life, photosynthetic bacteria, and Metazoa. Studies on the early evolution of life with fossil records began with discussions on the 1.9 Ga fossil bacteria. Currently, oldest fossil records are traced back to 3.5 Ga. Recent controversies on these fossils have been made on the basis of several geochemical approaches. Some interesting attempts for the detection of the photosynthetic activity from sedimentological setting of the strata-containing bacteria fossils should be noted. Ediacaran fossils are regarded as ancestors of Metazoa, but some researches proposed that apparent difference exists between those fossils and Metazoa. Recent discoveries of well-preserved three-dimensional Ediacaran fossils will provide insight into the evaluation of such unique organisms.(Keyword) fossil, Archean, microbial mat, Ediacara fauna, geology, Earth history
先カンブリア時代の地層に残された"化石"とその解読
川上紳一1,東條文治2
1 岐阜大学教育学部.
2 名古屋大学大学院環境学研究科.
要旨
生命の初期進化の重要な出来事はすべて先カンブリア時代に起こったといってよい.生命進化を探る上で重要な物証となる先カンブリア時代の化石記録については,その数や質的にも制約が多かった.地球化学的な分析の導入や,保存状態の良い化石が複数見つかる中で,生命の初期進化に関する研究は単純な形の比較を超えた新しい角度からの議論に進みつつある.初期生命進化に対する化石記録からの最新のアプローチを簡単に紹介したい. 生命の初期進化に関して注目される重要な出来事は,生命活動の開始,光合成細菌の出現,多細胞動物の出現などである.初期生命の化石記録の研究は19億年前のバクテリア形態に似た化石の記載から始まったが,その後バクテリア状の化石の発見は35億年前にまでさかのぼることになる.さらに地球化学的な手法によって生命活動の評価が議論されるようになってきた.光合成活動を確認することは難しいが,産出する地層の情報から評価する動きが出ている.多細胞動物の出現については,約5億5千万年前ごろから豊富に産出するエディアカラ生物群が有力な化石記録とされてきた.しかし,これが現生の多細胞動物ではないという議論もある.三次元形態保存の良い化石にもとづいてエディアカラ生物群化石がなにものであったのか探る研究が進んでいる.(キーワード)
化石、太古代、バイオマット、エディアカラ生物群、地質、地球史
1.はじめに
古い時代の地層から生命の起源と初期進化を探る試みは,スタンレイ・ミラーが生命の起源を実験的に調べる研究を行ってまもない1950年代から始まった.それから半世紀が経過して,膨大な研究が行われ初期地球の生命像に関する知見が急激に増えている[1,2,3].これまでに報告されている先カンブリア時代の化石にはバクテリア様の細胞化石,比較的大型で有機質のフィラメント状化石,多細胞動物化石がある[4].これらはいずれも生物進化における大きな躍進を示すものとして注目されてきた.しかし,証拠として提示された化石や地球化学的分析データには対立する解釈が提示され,論争が続いているものも多い.まず最大の批判は,細胞状の化石とされているものが,本当に生物の細胞が化石化したものなのか,それとも無機的にできた微小な物質が細胞状の生物に似ているに過ぎないのかという点である.この論争は,形態の問題であるが,形が単純であるだけに簡単には決着がつかないものである.こうした現状から,生物の起源と初期進化を研究する地質学者たちは,微生物様の化石の発見を中心に据えた研究から一歩踏み込んで,それらが発見された地層の堆積環境を基に,それがどのような生物だったのかを考察する研究へと向かっているが,こうした研究で得られたデータの解釈には,微生物学や微生物生態学といった異分野からの視点が重要である. 一方,先カンブリア時代の化石記録のなかで近年注目されているものにエディアカラ生物群に代表される初期多細胞動物がある.エディアカラ生物群化石の分類学的位置づけについても論争が続いているが,近年保存のよい化石産地が発見され,新たな研究の展開がみられるようになった.本論では,こうした先カンブリア時代の化石記録に基づく生物進化の研究の現状と今後の課題について述べる.
2.太古代の生命活動の痕跡
(1)研究小史地球の歴史を記録した堆積岩を調べていくと,カンブリア紀より古い時代の地層からはほとんどまったく化石が見つからなくなる.カンブリア紀以降の化石のほとんどは,貝化石や動物骨格化石などの石灰質の硬骨格が化石として残りやすい.しかし,地球の歴史をさらに遡った先カンブリア時代には生物はいなかったのか,それとも化石として残らなかったのか.カンブリア紀以降の化石の多くは,大型の動物化石や植物化石である.生物の系統分類学によると,生物界は大きくバクテリアなどの原核生物(モネラ)界,単細胞真核生物(プロチスタ)界,菌界,動物界,植物界に分けられる.カンブリア紀以降の地層から発見される多くの化石は,動物,植物化石なので,先カンブリア時代にも原核生物(バクテリア)や単細胞真核生物が繁栄していた可能性がある.これらの生物は微小なので地層中で化石として残りにくいことや,たとえ地層中に含まれていても発見される可能性は極めて低いと考えられてきた.すなわち,先カンブリア時代は地球が誕生した46億年前から5億4500万年前までの地球史の90%における生物進化は深い闇の中にあった.
1954年アメリカの地質学者E.S.BarghoornとS.A.Tylerは,アメリカとカナダの国境の五大湖の縁に露出する19億年前のガンフリント縞状鉄鉱床に含まれる黒色のチャートを調べ,バクテリアの形態とよく似たものが含まれていることを発見し,科学雑誌「サイエンス」に発表した[5].これは生物の初期進化の証拠として認識されるには,きちんと記載された論文として発表される必要があった.
それから10年の歳月が流れ,BarghoornとTylerが論文を仕上げる前に,P.Cloudがガンフリント縞状鉄鉱床から産出される化石の地球史的重要性を認識し,同じく雑誌「サイエンス」に発表しようとした[6].この動きを知ったBarghoornは若い大学院生J.W.Schopfを仲間に取り込んで,1954年の化石の記載論文を完成させた[7].こうした1960年代半ばになって,先カンブリア時代の黒色をしたチャートからバクテリア様の化石を見つける研究に火がつけられた[8,9].
やがて,オーストラリア,南アフリカ,グリーンランド,カナダなどでバクテリア様化石の発見論文が相次いで発表された.しかし,それらの中にはどう見ても生命化石とは考えられないようなものも含まれていた.Schopfは,こうした論文を一つひとつ詳細に検討し,それらが化石なのかにせものなのかを判定したレビュー論文を発表していった[1,2].また,古生物学的研究だけでなく,同位体地球化学,有機地球化学や地質学など周辺分野と連携した学際的機運が高まっていった.
1980年代半ばには,西オーストラリアのピルバラで約35億年前のチャート層からバクテリア様の化石が発見された[10].また,この地域では,ドーム状をした縞状堆積岩も発見され,微生物マットがつくったストロマトライトであると解釈された[11,12].これらの発見は,35億年前に光合成をする原核微生物であるシアノバクテリアが当時すでに繁殖していたことを示唆するものと考えられた.
(2)最古の生命活動の痕跡をめぐる論争
西グリーンランドのイスア地域は,世界最古の地層が露出する地域として1970年代から注目されてきた.ここでは,礫岩,砂岩,炭酸塩岩,縞状鉄鉱床などが露出しており,当時すでに陸と海が存在していたことを証明した.イスア地域の地層ができた時代にすでに生命は発生したのかにも当然注目が集まった.しかし,この地域は650℃に達する熱変成作用を受けているため微生物様の化石の探索や有機物の検出には困難がともなった.Schidrowskiは,この地域から採集されたグラファイトの炭素の同位体比を測定した[13].得られた結果-10〜-15艪ニいう値であり,分析した試料が生物起源の有機物に由来するものか,無機的物質なのかは明確にできなかった.しかし彼は,試料はそもそもシアノバクテリアの光合成でできたものが熱変成作用を受けたものであると解釈し,38億年前にすでに光合成を行って酸素を発生する生物が存在したと主張した.
1996年にMojzsis et al.は,イスア地域より年代が古いと考えられる西グリーンランドのアキリア島でアパタイトという鉱物を採集し,鉱物の中に含まれる微小なグラファイトの炭素同位体比を測定する技術を開発し,分析データを発表した[14].この研究で得られた炭素同位体比は-30〜-50艪ニいった値を示した.彼らは変成作用による同位体比の変化の可能性も検討し,それらが生物起源のグラファイトであると結論づけた.ところが,その後,彼らが試料を採集した地域の地層の解釈を巡って反論が持ち上がった.問題の地層はきれいな層構造が認められていたが,地質調査と岩石の微量元素組成を分析した研究者は,それらが堆積岩ではなく玄武岩などの火成岩が変成作用を受けて片麻岩という岩石になったものであると主張した[15].もし,問題の地層が火成岩起源であるとしたら,その岩石に含まれるグラファイトは生物起源である可能性はなくなる.現在,アキリア島の研究については,岩石の成因そのものが問題となっており,地層が堆積岩か火成岩かといった初歩的なところから再検討をしなくてはならなくなった.
一方,イスア地域については,Shidrowskiの研究[13]のあとに,炭素同位体比の追試がなされている.Rosingは,イスア地域の岩石に含まれる微小なグラファイトの炭素同位体比を分析し,生物起源であると論じている[16].Ueno et al.も同様にこの地域の岩石中のグラファイトの炭素同位体比を測定しているが,彼らは変成作用の度合いが低いほど炭素同位体比が負の値になることから,初期の炭素同位体比は生物起源の値をとっていたという評価がなされた[17].
(3)35億年前の生命化石を巡るその後の論争
1993年にSchopfはさらにピルバラで採集した岩石からシアノバクテリア様の化石を発見し,それらを詳細に記載した論文を発表した[18].それらが本当にシアノバクテリアの化石といってよいかは,あまり形態的特徴のない現生のシアノバクテリアの形と化石の形の比較によるもので,それらが化石であるかどうかの認定は主観がはいりやすいという欠点があった.
1990年代後半になると,シアノバクテリア様の"化石"をミクロスケールで研究する手法が確立された.まず,"化石"にレーザーを照射して含まれる炭素を取りだし,炭素同位体比を測定できるようになった[19,20]."化石"は岩石の変成作用を受けてグラファイトになっているが,一部は変質した有機物(ケロジェン)として残っている可能性がある.いずれにしても炭素同位体比の値から生物起源なのか,非生物起源なのか議論できるようになった.得られたデータは,生物がつくった有機物と重なる同位体比をもっていた.しかし,炭素同位体比は変成作用時に同位体交換反応が起こって変化していて,形成時の情報をとどめていないのではないかという批判がつきまとった.
さらに,こうした研究と研究手法への検証が,1996年に発表された火星起源いん石中の生命活動の痕跡にまつわる世界中の研究者によって注目された.その結果,1993年に発表されたSchopfの論文[18]のように形態比較に基づく生命化石の認定に落とし穴があることが指摘された.
まず,形態の比較について異論があり,これまでシアノバクテリアと類似した形態といわれていたものも,よく見ると分岐構造が見えたりして,必ずしも形がぴったり一致しているというわけではないという批判がでた[21].さらに化石と似たような形態をした物質が熱水溶液中で合成できたとする研究発表もだされた[22].
Schopfらは,これまでに先カンブリア時代の岩石から報告されているバクテリア様化石について,ラマンスペクトルを測定して,生物起源であることを裏づけようとした[23].しかし,ラマンスペクトルは,炭素原子の結合状態を調べるもので,たとえ"化石"を形づくっているものがケロジェンでできていることが確かめられたとしても,それが生物起源の証拠にはならないという批判がでている.このように見てみると,先カンブリア時代,とりわけ太古代の岩石からバクテリア様の化石を発見したとしても,化石ではないという反論をしりぞけるための分析手法,判定基準が明確ではなく,論争がさらに拡散していくことになっている.
そこで,地質学者は論点を変えて,微生物化石や生物がつくったマットがどのような堆積環境のもとで形成されたかに論点を移している.まず,Ueno et al.は,ピルバラ地域の地質構造を詳しく解析し,当時そこが中央海嶺近くの熱水噴出孔付近であり,玄武岩質の地殻を覆うチャート層は熱水沈殿物であることを明らかにした[20].さらに,その水深が1500mという見積もりがだされたため,発見された微生物様化石は,光の届かない深海底で生息する光合成細菌である可能性が示唆された.Rassmusonも,硫化物鉱床から採集された岩石に微生物様化石を発見し,それが深海底に生息していた生物であると主張している[24].
こうした知見は明らかにShopfの結論[18]と矛盾している.Rassmusonは,当時浅海域から深海底まで生物の生息環境は広がりを見せていたという解釈を行った[24].しかし,浅海域で生息していたとされる証拠として注目されたピルバラ地域の最古のストロマトライトについては,生物起源であるという解釈に反論が提示されており[25],浅海域に生態系が形成されていたという解釈には否定的な見方もある.これまでに得られた情報を総合すると,ピルバラ地域の地層からは光合成を行って酸素を発生する生物が当時出現していたかどうかについては,有力な証拠に乏しくなったようにみえる.
(4)35億年前の光合成細菌のマットとは
南アフリカの35億年前の有機質チャートの堆積環境から,当時浅海域に光合成細菌(光合成を行うが酸素を発生しない原核生物)が生息していたとされる論文が最近発表された[26].彼らは,バック・リーフ・チャートから微生物様の化石や,微生物の集合体がぬるぬるとした有機質の皮膜(バイオマットと呼ばれている)をつくっていた証拠を多数発見した.彼らはこうした地層がどのような環境で形成されたかを論点の中心にすえた.そしてそのために必要なデータを広域的な地質調査によって集めた.この地層は,太陽光の差し込む浅い海洋で形成されたものであり,その地層の下位には海水が蒸発してできた蒸発岩の地層が堆積している.一方,上位にはより沖合で堆積した縞状鉄鉱床が堆積している.
こうした地層の重なりからどのようなことがいえるのか.まず,一連の地層が浅い海洋で形成したものであり,沖合で縞状鉄鉱床が堆積していることは,当時海水が成層構造しており,深海の鉄に富んだ海水と表層海水が混ざり合って鉄が沈殿したものと考えられる.ただし,この縞状鉄鉱床は鉄炭酸塩鉱物(FeCO3)であり,一般的な縞状鉄鉱床のように酸化鉄鉱物ではないことに注意する必要がある.つまり,鉄が酸化物ではなく炭酸塩鉱物として堆積していることは,問題のバイオマットは酸素を発生する光合成微生物(シアノバクテリア)が構築したものではないことになる.
また,この地域には熱水活動の証拠は認められなかったため,有機質のチャートは熱水性のものではなく,海水から析出した鉱物からできていると解釈された.これは,この地域の地層が浅海域でできたとする解釈と矛盾しない.
では,バイオマットの正体はどのように考えたらよいのだろうか.バイオマットをつくる微生物は,(1)光合成を行わず酸化還元反応でエネルギーを獲得する化学合成細菌,(2)光合成を行うが酸素を発生しない微生物(専門家はこれを光合成細菌という),(3)酸素を発生するシアノバクテリアや藻類の3つのタイプがある. (3)は縞状鉄鉱床の構成鉱物が酸化物でないことから除外された.(1)は温泉や深海熱水噴出孔のように,酸化還元状態の異なる物質が出会う環境でバイオマットを作るものである.(2)は湖や温泉など,太陽光が利用できるさまざまな環境に生息している.いずれもバック・リーフ・チャートの有機物を作り出した微生物の候補と考えられるが,太陽光が差し込むほどの浅海であることから,TiceとLoweは(2)の解釈を採用したということができるだろう.
したがって,35億年前に光合成細菌が生息していたという主張の根拠はあくまでも地質学的なものであり,しっかりした微生物学的な根拠があるわけではないことに注意する必要がある.彼らの議論で欠落している点は,現在地球に棲息している光合成細菌は,硫化水素を酸化して独立栄養的にエネルギーを獲得するか,周囲の有機物を取り込んで従属栄養的にエネルギーを獲得していることである.もし,35億年前のバイオマットが光合成細菌が構築したものだとすると,硫化水素が存在するような場所だったことになる.こうした状況では鉄は炭酸塩鉱物ではなく硫化物からなる縞状鉄鉱床を形成することになる.
3.多細胞動物の出現
(1)多細胞動物の出現という出来事生命の誕生と,光合成生物の登場,真核生物の出現に続いておこる生命の歴史での重要な出来事に,多細胞動物の出現がある.多細胞動物の出現は生命進化において画期的な出来事であったということに異論を持つ人は少ないだろう.多細胞化が起こると当然の帰結として生物は大型化し,複雑化することが予想できる.そして細胞が分化し,それぞれに役割分担を振り分けることによってさまざまな機能を獲得することができる.環境への適応能力や捕食といった高次の生態系システムの構築など,多細胞動物の出現によって生命現象の描像は決定的にそれまでのものと違うものになった.
最初の多細胞動物がどのようなものであったか考える上で化石記録は重要な物証となる.近年,原生代後期の地層から多細胞動物と考えられるさまざまな化石が新たに発見され,化石記録からの研究は意欲的に進んでいる.化石記録からは約6〜10億年前ごろの多細胞動物の化石とされるものがいくつも報告されているが,必ずしも確実なものではない.複数の同様の形態の化石の産出によって信頼性の高い議論ができるようになるのは中国のドウシャントウの微化石(約5億7000万年前),世界各地から見つかるエディアカラ生物群化石(5億6500万年前〜5億4300万年前)である.エディアカラ生物群化石は多様な化石を含み,数mにおよぶ巨大な個体も産出し,発見当初は環形動物や刺胞動物といった現生につながる多細胞動物を含むと考えられたため,最古の多細胞動物群として注目されてきた.現生の多細胞動物の系統を考える上でも,多細胞動物出現初期の生物界の姿を考える上でも重要な物証として研究されている.
(2)エディアカラ生物群化石の研究史
エディアカラ生物群化石が最初に発見されたのは,1908年アフリカ,ナミビアにおいてであるが,1946年にSpriggが南オーストラリアのフリンダースレンジ,エディアカラの丘で発見した生物群化石(それらは580から543Ma)が有名となり,エディアカラ生物群化石と呼ばれるようになった[4].その後,世界各地のこの時代の地層から同様の化石群が発見されている.Glaessnerはエディアカラ生物群化石(特に先カンブリア時代の後期に保存されている)の中に刺胞動物,節足動物,環形動物などが含まれると考えたが[27],形態が原始的であるためにその根拠に乏しいものであった.Dickinsoniaは,キルト構造と呼ばれる体節様の構造を持ち,多毛類に似ていることから環形動物と考えられ,Sprigginaは同様のキルト構造を持つが節足動物と考えられた(Fig.1).Ediacaria(Fig.2)などはクラゲ様の同心円状の構造から,刺胞動物とされたが,これらの化石はCharniodiscus(Fig.1)の下部に位置する固定円盤の部分であることがのちに明らかになっている.
Seilacherは,エディアカラ生物群の大部分を占めるDickinsoniaなどの,キルト構造を持つ生物たち(Fig.1)は,多細胞動物ではないとした研究を発表した[28].その理由は,これらが大きさに比べて厚さが非常に薄いこと,細長いキルト構造が非常に密に繰り返されること,さらにキルト構造は一見すると左右対称に見えるが左右の構造は正中線にそって互い違いに接合することである.この論争は現在も決着がついていない.
その一方で,エディアカラ生物群の中に現在の多細胞動物との関係が有力な化石もある.Kimberella(Fig.2)がそれで,より立体的であること,左右相称的であること,フリル状の構造などの類似性から軟体動物の祖先ではないかと考えられている[29].しかし,エディアカラ生物群において産出量,種類数からいっても,Seilacherが多細胞動物ではないと主張しているキルト構造を持つタイプの化石が多数を占めている.エディアカラ生物群化石の研究が進むと,現在の多細胞動物につながらない当時の独特な生物群の繁栄の陰でわたしたちの祖先はひっそりと生活していたということになるかもしれない.とはいえ,エディアカラ生物の形態はその後の時代のバージェス動物群などと比較しても,明らかに形態が単純なものが多く,現生生物の形態との一次近似的な類似性に基づいた議論で,その系統学的分類学的位置づけを結論付けるには限界がある.エディアカラ生物群を現生生物との系統・分類学的な研究と直接結びつけるのではなく,この時代に繁栄したエディアカラ生物はどんな生き物であったのか,その生き様を理解しようという試みが増えてきている.
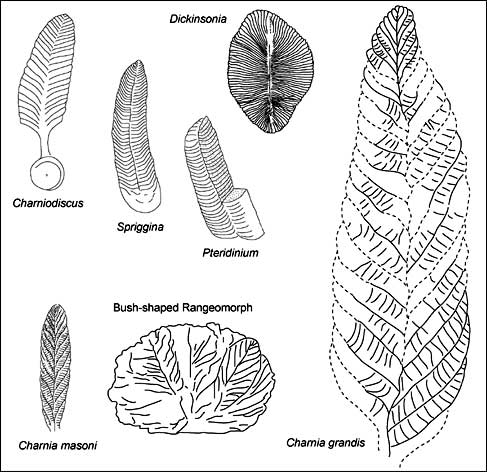

カナダのニューファウンドランドで三次元形態を保存したエディアカラ生物群化石が発見され[30],オーストラリア,ナミビア,ロシアなどの豊富な化石群とあわせて,エディアカラ化石群を構成する化石の形態の変化を時代を追って検討できるようになってきた.このなかで, BrasierとAntcliffeは,キルト構造をもつ化石グループの形態を詳細に観察すると,これらの形態の基本的単位が認定できると主張した[31].そして,形態の類似性から基本的な単位としてCharnia masoniを認定すると,Charnia grandisは巨大化したもの,Bush-shaped Rangeomorphは群体を形成したもの,などと考えることができるというのである(Fig.1).Charnia grandisの一部が再びCharnia masoniに似た形態をしていることから,これらを一連の再生産のサイクルである可能性も指摘している.
また,BrasierとAntcliffeは,各産地から産出する化石形態から進化的な傾向を見ると,キルト構造がより細かく分岐しないものが多くを占めるようになるため,キルト構造のフラクタル状の分岐が減少しシンプルな形に向かう進化傾向がみられると主張している[31].これらの見出した傾向について今後検証する必要はあるが,エディアカラ生物,そのものの生き様,時代変化を理解するという研究として新しい試みである.
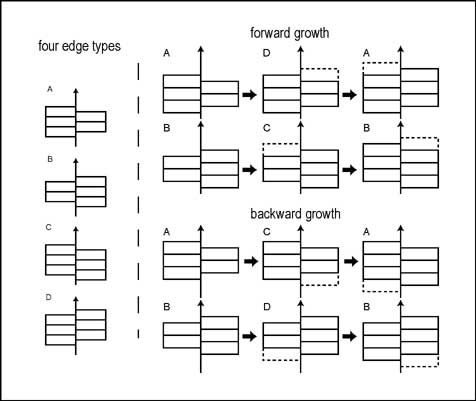
東條らは,エディアカラ生物群化石の中にキルト構造からを持つ形態が多く含まれていることに注目した.重要な特徴として接合線に沿って,互い違いに接合する点が挙げられる.この互い違いの構造は非常に正確な構造であり乱れが見られないことから,キルト構造の肋は正中線の端で付加されると考えた.キルト構造については,成長の様式によって,化石がもつ端のキルト構造のパターンはことなるため,産出する化石の個体群から成長様式を推定することができると考えた(Fig.3)[32].
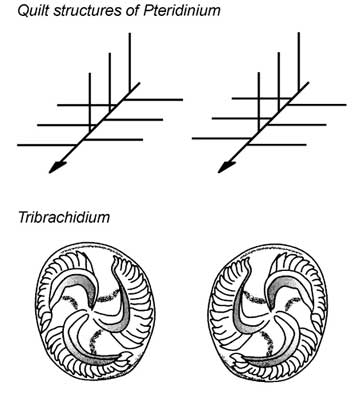
また,エディアカラ生物群の化石の中に,左右非相称な形態で右手系,左手系に対応する鏡像体を持つものが存在することを指摘している[32].Pteridiniumのようなキルト構造が接合軸において互い違いに接合するもの,さらにTribrachidiumのように3回回転対称の形態で鏡像体をもつものなどである(Fig.4).このように左右相称性が基本的な形態において崩れている生物がしばしば見受けられる.これらの鏡像体における,右手系・左手系の構成比率や,左右非相称な生物が占める割合の時代変化などといった,まったく新しい視点からエディアカラ生物群化石を考えようと試みている.
多細胞動物出現の時期とその原因についてはも多細胞動物出現の時期とその原因についてはもう一つの問題となっている.真核生物の登場から多細胞動物の登場までには非常に長い時間がかかっているという印象を受ける.多細胞動物出現のメカニズムの完成にはそれだけの時間が必要だったということなのか,環境的要因が大きく影響したのかは大きな問題だ.また,この多細胞動物が登場したと考えられている6〜10億年前は地球は全球的な規模の氷河時代であったことも知られている.この氷河時代については「全球凍結仮説」として近年注目を集めているが[33,34],この氷河時代が多細胞動物の出現に大きな影響を与えたと考える研究者は少なくない.Kirschvinkは,氷河時代の後,光合成活動が活発になり酸素分圧が上昇したため,多細胞動物が出現したという説を出している[35].Hoffmanも氷河時代に生物が複数の避難地に隔離され進化速度が上昇したという仮説[36]に言及している. 現在の段階ではこれらの考えに具体的な物証を与えることは難しいが,化石記録などによって初期の多
細胞動物がどのようなものであったかが明らかになってくると,その時期の必然性について大きな示唆があたえられるかもしれない.
4.結論
化石記録から初期生命に迫る研究は,現生生物との形態の比較からはじまり,さまざまな分析手法の導入を経て,十分な評価を試みてきた.地球化学的な手法や,地質学的な検討も含めてさまざまな試みが行われてきたが,本当に生命活動の証拠であるか決定的な評価法は定まっていない.光合成活動についての検討では化石が産出する地層から当時の環境を復元し,状況証拠から迫る試みが行われているが,エネルギー代謝システムの生成物の痕跡について十分に検討する必要がある.多細胞動物の出現については,初期の原始的形態の生物を現生生物の形態と比較して分類・系統を議論することには限界がある.分類・系統学的な着地点の模索ではなく,当時の生物の生態や生活様式に関する理解をまず豊富にするという方向へ研究は進みそうだ.先カンブリア代の化石記録はわずかで,現状では初期生命の謎の議論に十分な決着が与えられてない点も多い.新しい分析や新しい化石記録の発見が研究を大きく前進させる分野だけに論点を整理して,現在の議論の限界を十分理解しておきたいところだ.文献
1. Schopf, J. W. Earth's Earliest Biosphere. Its Origin and Evolution. Princeton Univ. Press, (1983).2. Schopf, J. W. and Clein, C. The Proterozoic Bioshpre. Cambridge Univ. Press, (1992).
3. Bengtson, S. Early life on Earth. Columbia University Press, (1994).
4. 大野照文,先カンブリア時代からカンブリア紀の生命の歴史,シリーズ進化学1−第3章,岩波書店,2004.
5. Tyler, S. A. and Barghoorn, E. S. Occurrence of structurally preserved planets in pre-Cambrian rocks of the Canadian shield. Science, 119, 606-608 (1954).
6. Cloud, P. Significance of the Gunflint (Precambrian) microflora. Science, 148, 27-45 (1965).
7. Barghoorn, E. S., and Tyler, S. A. Microorganisms from the Gunflint chert. Science, 147, 563-577 (1965).
8. Barghoorn, E. S., Meinschein, W. G. and Schopf, J. W. Paleobiology of a Precambrian shale. Science, 148, 461-472 (1965).
9. ウイリアム・ショップ,失われた化石記録,講談社現代新書,1998.
10. Schopf, J. W. and Packer, B. M. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science, 237, 70-73 (1987).
11. Walter, M. R., Buick, R. and Dunlop, J. R. S. Stromatolites 3,400-3,500 Myr old from the North Pole area, Western Australia. Nature, 284, 443-445 (1980).
12. Lowe, D. R. Stromtolites 3,400-Myr old from the Archaean of Western Australia. Nature, 284,441-443 (1980).
13. Schidrowski, M. A 3800-million year old isotopic record of life from carbon in sedimentary rocks. Nature, 333, 313-318 (1988).
14. Mojzsis, S. J., Arrhenius, G., McKeegan, K. D., Harrison, T. M., Nutman, A. P. and Friend, C. R. L. Evidence for life on Earth before 3,800 million years ago. Nature, 384, 55-59 (1996).
15. Fedo, C. M. and Whitehouse, M. J. Metasomatic origin of quartz-pyroxene rock, Akilia, Greenland, and implications for Earth's earliest life. Science, 296, 1448-1452 (2002).
16. Rosing, M. T. 13C-depleted carbon microparticles in >3,700-Ma sea-floor sedimentary rocks from West Greenland. Science, 283, 674-676 (1999).
17. Ueno, Y., Yurimoto, H., Yoshioka, H., Komiya, T. and Maruyama, S. Ion microprobe analysis of graphite from ca. 3.8 Ga metasediments, Isua supracrustal belt, West Greenland: Relationship between metamorphism and carbon isotopic composition. Geochim. Cosmochim. Acta, 66, 1257-1268 (2002).
18. Schopf, J. W. Microfossils of the early Archean Apex chert: new evidence of the antiquity of life. Science, 260, 640-646 (1993).
19. House, C. H., Schopf, J. W., McKeegan, K. D., Coath, C. D., Harrison, T. M. and Stetter, K. O. Carbon isotopic composition of individual Precambrian microfossils. Geology, 28, 707-710 (2000).
20. Ueno, Y., Isozaki, Y., Yurimoto, H. and Maruyama, S. Carbon isotopic signatures of individual Archean microfossils (?) from Western Australia. Internal Geology Reviews, 43, 196-212 (2001).
21. Brasier, M. D., Green, O. R., Jephcoat, A. P., Kleppe, A. K., Van Kranendonk, M. J., Lindsay, J. F., Steele, A. and Grassineau, N. V. Questioning the evidence for Earth's oldest fossils. Nature, 76-81 (2002).
22. Garcia-Ruiz, J. M., Hyde, S. T., Carnerup, A. M., Christy, A. G., Van Kranendonk, M. J. and Welham, N. J. Self-assembled silica-carbonate structures and detection of ancient microfossils. Science, 302, 1194-1197 (2003).
23. Schopf, J. W., Kudryavtsev, A. B., Agresti, D. G., Wdowiak, T. J. and Czaja, A. D. Laser-Raman imagery of Earth's earliest fossils. Nature, 416, 73-76 (2002).
24. Rasmussen, B. Filamentous microfossils in a 3,235-million-year-old volcanogenic massive sulphide deposit. Nature, 405, 676-679 (2000).
25. Lowe, D. R. Abiological of described stromatolites older than 3.2Ga. Geology, 22, 387-390 (1994).
26. Tice, M. M. and Lowe, D. R. Photosynthetic microbial mats in the 3,416-Myr-old ocean. Nature, 431, 549-552 (2004).
27. Glaessner, M. F. The dawn of animal life, Cambridge Univ. Press, (1984).
28. Seilacher, A. J. Vendobionta and Psammocorallia: lost constructions of Precambrian evolution. Geol. Soc. London, 149, 607-613 (1992).
29. Fedonkin, M. A. The origin of the Metazoa in the light of the Proterozoic fossil record. Paleontological Research, 7, 9-41 (2003).
30. Narbonne, G. M. Modular Construction of Early Ediacaran complex life forms. Science, 305, 1141-1144 (2004).
31. Brasier, M. and Antcliffe, J. Decording the Ediacaran enigma, Science, 305, 1115-1117 (2004).
32. 東條文治・斎藤良・川上紳一・大野照文,エディアカラ生物群にみられるキルト構造の理論形態,月刊地球,26,177-182 (2004).
33. Hoffman, P. F., Kaufman, A. J., Halverson, G. P. and Schrag, D. P. A Neoproterozoic snowball Earth. Science, 281, 1342-1346 (1998).
34. 川上紳一,全地球凍結,集英社新書,2004.
35. Kirschvink, J. L., Gaidos, E. J., Bertani, L. E., Beukes, N. J., Gutzmer, J., Maepa, L. N. and Steinberger, R. E. Paleoproterozoic snowball Earth: extreme climatic and geochemical global change and its biological consequences. PNAS, 97(4), 1400-1405 (2000).
36. Hoffman, P. F. Snowball Earth. Scientific American, Jan, 68-75 (2000).