ORIGIN OF PHOTOSYNTHESIS AND ITS EVOLUTIONARY PROCESS
Satoshi Hanada
Institute for Biological Resources and Functions
National Institute of Advanced Industrial Science and Technology (AIST)
Tsukuba Central 6, 1-1-1 Higashi, Tsukuba 305-8566, Japan.
Tel: +81-29-861-6590ĄĄFax: +81-29-861-6587
E-mail
(Received 18 August 2004 Accepted 12 November 2004)
(Abstract)
Cyanobacteria can oxidize water using light energy, and which causes evolving oxygen molecules. In the history of life on the Earth, the origin of cyanobacteria is not so old, and anoxygenic photosynthesis has been established prior to the first appearance of oxygenic photosynthesis. Such phototrophs that photosynthesize without oxygen production still survive up to now and frequently found in various environments. Anoxygenic photosynthetic bacteria are taxonomically divided into four groups: filamentous anoxygenic phototrophic bacteria (FAPB), purple bacteria (PB), green sulfur bacteria (GSB), and heliobacteria (HB). Cyanobacteria are known to use two different types of photopigment-containing protein complex, i.e., photosystem I (PS I) and photosystem II (PS II), in their oxygenic photosynthesis, whereas anoxygenic phototrophs possess only one of two photosystem-types. GSB and HB contain a PS I-type photosystem in their photosynthesis, while FAPB and PB contain a PS II-type photosystem. The phylogenetic analysis based on 16S rRNA sequence and comparison of the photochemical structure among the phototrophic bacteria suggested that the origin of anoxygenic photosynthesis is much older than that of oxygenic photosynthesis and a photosynthetic core complex in the first phototroph appeared to be similar to a PS II-type photosystem. In the evolutionary pathway of photosynthesis, a PS II-type phototroph emerged first, and then a PS I-type phototroph came out. Oxygenic photosynthesis that appeared subsequently to anoxygenic photosynthesis originated from genetic incorporation of these two different types of photosystems.(Keyword) photosynthesis, anoxygenic photosynthetic bacteria, cyanobacteria, oxygen evolution
ļũđįĀŪĪÎ―Ðļ―ĪČŋĘē―
ēÖÅÄĄĄÃŌĄĄĄĘŧšķČĩŧ―ŅÁíđįļĶĩæ―ęĄĶĀļĘŠĩĄĮ―đĐģØļĶĩæÉôĖįĄË ÆČÎĐđÔĀŊËĄŋÍĄĄŧšķČĩŧ―ŅÁíđįļĶĩæ―ęĄĘAISTĄËĄĄĀļĘŠĩĄĮ―đĐģØļĶĩæÉôĖį
ĒĐ305-8566ĄĄ°ņūëļĐĪÄĪŊĪÐŧÔÅė1-1-1ĄĄĪÄĪŊĪÐÂč6ŧöķČ―ę
1. ĪÏĪļĪáĪË
ļ―šßĪÎÂįĩĪĪÎĢĩĘŽĪÎĢąĪōĀęĪáĪëŧĀÁĮĪÏĄĪžįĪËĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪČļÆĪÐĪėĪëŧĀÁĮČŊĀļ·ŋĪÎļũđįĀŪšŲķÝĪËĪčĪęĀļŧšĪĩĪėĪŋĪâĪÎĪĮĪĒĪëĄĨĪģĪÎĪčĪĶĪĘÂįÎĖĪÎŧĀÁĮĪÎČŊĀļĪŽÃÏĩåīÄķĪËÍŋĪĻĪŋÂŋÂįĪĘĪëąÆķÁĄĪĀļĘŠÁīÂÎĪËÂÐĪđĪëĪ―ĪÎļųšáĪËīØĪ·ĪÆĪÏĄĪšĢĪĩĪéļėĪëĪŲĪŊĪâĪĘĪĪĪŽĄĪĪģĪÎĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪËĪčĪëŧĀÁĮČŊĀļĪÏļũĨĻĨÍĨëĨŪĄžĪōÍŅĪĪĪŋŋåĪÎĘŽēōĄĘŧĀē―ĄËĪŽļķ°øĪČĪĘĪÃĪÆĪĪĪëĄĨĪÄĪÞĪęĄĪÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆĪÎŋåĪŦĪéÅÅŧŌĪō°úĪČīĪŊĪŋĪáĄĪÉûŧšĘŠĄĘÏ·ĮŅĘŠĄËĪČĪ·ĪÆĘŽŧŌūõŧĀÁĮĪŽĀļĀŪĪ·ĪÆĪ·ĪÞĪĶĪïĪąĪĮĪĒĪëĄĨ ļũđįĀŪĩĄĮ―ĪÎŋĘē―ēáÄøĪĮĪÏĄĪŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎ―Ðļ―ĪËĀčÎĐĪÃĪÆŧĀÁĮĪÎČŊĀļĪōČžĪïĪĘĪĪĨŋĨĪĨŨĪÎļũđįĀŪĄĘČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĄËĪŽÂļšßĪ·ĪŋĪâĪÎĪČđÍĪĻĪéĪėĪÆĪĪĪëĄĨĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪŽÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆŋåĪōÍŅĪĪĪëĪģĪČĪŽ―ÐÍčĪëĪÎĪËÂÐĪ·ĄĪĪģĪėĪéČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏŋåĪōķĄÍŋÂÎĪČĪđĪëĪģĪČĪŽ―ÐÍčĪšĄĪÂåĪïĪęĪËÎēē―ŋåÁĮÅųĪÎīÔļĩÅŠÎēēŦē―đįĘŠĪäŋåÁĮĄĘĪÞĪŋĪÏÍĩĄŧĀĄËĪĘĪÉĪōŧČĪÃĪÆļũđįĀŪĪōĪđĪëĄĨĪģĪÎĪčĪĶĪĘČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪōđÔĪĶšŲķÝĪâļ―ÂļĪ·ĪÆĪŠĪęĄĪĪ―ĪėĪéĪÏÂįĪĪŊ°ĘēžĪÎĢīĪÄĪÎĨ°Ĩ륞ĨŨĪËĘŽĪąĪéĪėĪÆĪĪĪë Ą§ đČŋ§ļũđįĀŪšŲķÝĄĘPurple bacteriaĄËĄĪÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĄĘFilamentous anoxygenic phototrophic bacteriaĄËĄĪÎÐŋ§ÎēēŦšŲķÝĄĘGreen sulfur bacteriaĄËĄĪĨØĨęĨŠĨÐĨŊĨÆĨęĨĒĄĘHeliobacteriaĄËĄĨ ĪģĪėĪéĢīžïÎāĪÎČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏĪ―ĪėĪūĪė·ÏÅýŋĘē―ÅŠĪËÎĨĪėĪÆĪŠĪęĄĪĪÞĪŋĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪŦĪéĪâģÖĪŋĪÃĪÆĪĪĪëĄĨ šŲķÝĪηÏÅýĪËĪŠĪĪĪÆĄĪĄÖČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎĢīĨ°Ĩ륞ĨŨĪŽđĪŊĘŽŧķĪ·ĪÆĪĪĪëĪģĪČĄŨ ĪČ ĄÖŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎÂļšßĪŽĢąĨ°Ĩ륞ĨŨĪËļÂÄęĪĩĪėĪÆĪĪĪëĪŽŧķšßĪ·ĪÆĪĪĪëĪģĪČĄŨĪÏĄĪ1)ļũđįĀŪĩĄĮ―ĪÏĀļĖŋĪÎŋĘē―ĪÎĪīĪŊ―éīüĪËģÍÆĀĪĩĪėĪŋĪâĪÎĪĮĪĒĪëĪģĪČĄĪ ĢēĄËŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎĩŊļŧĪÏĪ―ĪėĪčĪęôĢĪŦĪËŋ·Ī·ĪŊĄĪĪ―ĪėĪÞĪĮĪÎļÅÃÏĩåĀļÂÖ·ÏĪĮĪÏļũ°ÍÂļ·ŋĀļĘŠ(phototrophs)ĪÏĪđĪŲĪÆČóŧĀÁĮČŊĀļ·ŋĪĮĪĒĪęĄĪĪ―ĪÎĪčĪĶĪĘŧþÂåĪÏĪŦĪĘĪęÄđĪŊÂģĪĪĪŋĪģĪČĪōķŊĪŊžĻšķĪđĪëĄĨËÜÏĀĀâĪĮĪÏĄĪšŲķÝĪËĪŠĪąĪëļũđįĀŪĪΚŽīīĪĮĪĒĪëļũē―ģØČŋąþÃæŋīĄĘËÜÏĀĪĮĪÏ ÃąĪËļũē―ģØ·ÏphotosystemĪČļÆĪÖĄËĪÎÂŋÍÍĀĪČĪ―ĪÎĩŊļķĄĪŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎģÍÆĀĪËŧęĪëŋĘē―ēáÄøĪËĪÄĪĪĪÆÏĀĪļĪëĪČĪČĪâĪËĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒ―Ðļ―°ĘÁ°ĄĘĘŽŧŌūõŧĀÁĮĪŽĪÛĪČĪóĪÉÂļšßĪ·ĪĘĪŦĪÃĪŋŧþÂåĄËĪÎČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪŽļÅÃÏĩåīÄķĪËÍŋĪĻĪŋąÆķÁĪäĪ―ĪÎĖōģäĪËĪÄĪĪĪÆđÍŧĄĪ·ĪŋĪĪĄĨ
ĢēĄĨļũđįĀŪšŲķÝĪÎĘŽÎā
ŋĒĘŠĪäÁôÎāĄĘÎÐÁôĪäđČÁôĄĪģėÁôĪĘĪÉĄËĪÏļũđįĀŪĪËČžĪĪŧĀÁĮĪōČŊĀļĪđĪëĄĨĪģĪÎŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÏšŲËĶÆâūŪīïīąĪĮĪĒĪëÍÕÎÐÂÎĄĘĨŊĨíĨíĨŨĨéĨđĨČĄËĪËĪčĪęđÔĪïĪėĪÆĪĪĪëĄĨļũđįĀŪĪōđÔĪĶŋŋģËĀļĘŠĪđĪŲĪÆĪΚŲËĶÆâĪËÂļšßĪđĪëÍÕÎÐÂÎĪÏĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĄĘÍõŋ§šŲķÝĄ§ĪŦĪÄĪÆĪÏÍõÁôĪČļÆĪÐĪėĪÆĪĪĪŋĄËĪČĪĪĪĶĨÐĨŊĨÆĨęĨĒĪÎÁÄĀčĪŽôĢĪŦĀÎĪËšŲËĶÆâķĶĀļĪ·ĪŋĪâĪÎĪĀĪČđÍĪĻĪéĪėĪÆĪĪĪë[1]ĄĨĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÏļķģËĀļĘŠĪÎÃæĪĮŧĀÁĮČŊĀļ·ŋļũđįĀŪĪōđÔĪĶÍĢ°ėĪÎĀļĘŠĪĮĪĒĪëĄĨšŲķÝĪÎÃæĪËĪÏĪģĪÎĪčĪĶĪĘļũđįĀŪĪČĪÏ°ÛĪĘĪęŧĀÁĮĪÎČŊĀļĪōČžĪïĪĘĪĪļũđįĀŪĄĪĪđĪĘĪïĪÁĄÖČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĄŨĪōđÔĪĶšŲķÝĪŽÂļšßĪ·ĪÆĪĪĪëĄĨĪģĪÎĪčĪĶĪĘČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏšŲķÝ·ÏÅýĪÎÃæĪËŧķšßĪ·ĪÆĪŠĪęĄĪ°ĘēžĪÎĢīĪÄĪÎĨ°Ĩ륞ĨŨĪËĘŽĪŦĪėĪÆĪĪĪë[2]ĄĨĢąĄËÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝ (Filamentous anoxygenic phototrophic bacteria, FAPB)Ą§ÂŋšŲËĶÁĄ°ÝūõĪηÁÂÖĪōŧýĪÄļũđįĀŪšŲķÝĪĮĪĒĪęĄĪēđĀôĪäēÏĀîĪËĀļÂĐĪ·ĪÆĪĪĪëĄĨĪÞĪŋĄĪģĪīßĪÎĨŋĨĪĨÉĨŨĄžĨëĪäđâąöĮŧÅŲĪÎąöļÐĪĮĪâĪ―ĪÎÂļšßĪŽģÎĮ§ĪĩĪėĪÆĪĪĪëĄĨļũđįĀŪĪËĪčĪëĀļ°éĪĀĪąĪĮĪÏĪĘĪŊĄĪÍĩĄĘŠĪČŧĀÁĮĪŽÂļšßĪđĪëīÄķĪĮĪÏŧĀÁĮļÆĩÛĪËĪčĪëĀļ°éĪâēÄĮ―ĪĮĪĒĪëĄĨļũđįĀŪŋ§ÁĮĪČĪ·ĪÆĨÐĨŊĨÆĨęĨŠĨŊĨíĨíĨÕĨĢĨë (Bchl) aĪōīÞĪߥĪļũĘá―ļĪÎĪŋĪáĪÎĘä―õŋ§ÁĮĪČĪ·ĪÆBchl c (ĪâĪÎĪËĪčĪęBchl dĪäe) ĪōđįĪïĪŧŧýĪÄĪâĪÎĪŽÂŋĪĪĄĨ·ÏÅýĪČĪ·ĪÆĪÏChloroflexiĖįĪË°Ī·ĪÆĪĪĪë[3, 4]ĄĨ ĢēĄËđČŋ§ļũđįĀŪšŲķÝĄĘPurple bacteriaĄËĄ§ČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎÃæĪĮšĮĪâžïŋôĪÎÂŋĪĪĘŽÎā·ēĪĮĪĒĪëĄĨēÏĀîĄĪļÐŋåĄĪģĪÍÎĄĪÅÚūíĪĘĪÉÍÍĄđĪĘīÄķÃæĪËļŦĪéĪėĪëĄĨŧĀÁĮĪËÂÐĪ·ĪÆđâĪĪīķžõĀĪōžĻĪđĪâĪÎĪâūŊĪĘĪŊĪĘĪĪĪŽĄĪđČŋ§ČóÎēēŦļũđįĀŪšŲķÝĄĘPurple nonsulfur bacteriaĄËĪË°ĪđĪëšŲķÝĪÎÂŋĪŊĪÏŧĀÁĮļÆĩÛĪĮĀļ°é―ÐÍčĪëĄĨļũđįĀŪŋ§ÁĮĪČĪ·ĪÆBchl aĪÞĪŋĪÏBchl bĪōŧýĪÃĪÆĪĪĪëĄĨĪģĪėĪéĨ°Ĩ륞ĨŨĪÏĪđĪŲĪÆProteobacteriaĖįĪË°Ī·ĪÆĪĪĪë[5, 6]ĄĨ ĢģĄËÎÐŋ§ÎēēŦšŲķÝĄĘGreen sulfur bacteriaĄËĄ§ŧĀÁĮĪËÂÐĪ·đâĪĪīķžõĀĪōŧýĪÄŲĻĀ·ųĩĪĀĪÎļũđįĀŪšŲķÝĪĮĪĒĪëĄĨģĪÍÎĄĶÎĶŋåÁÐĘýĪËļŦĪéĪėĪëĄĨīÞÍĪđĪëļũđįĀŪŋ§ÁĮĪÏĀčĪÎÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪČÎāŧũĪ·ĪÆĪŠĪęĄĪBchl aĪČļũĘá―ļĪÎĪŋĪáĪÎŋ§ÁĮĪČĪ·ĪÆBchl c (ĪäBchl dĄĪe) ĪōŧýĪÃĪÆĪĪĪëĄĨĪģĪėĪéĪÏÎÐŋ§ÎēēŦšŲķÝĪÎĪßĪĮđ―ĀŪĪĩĪėĪŋChlorobiĖįĪČļÆĪÐĪėĪë·ÏÅýĘŽÎāĖįĪË°Ī·ĪÆĪĪĪë[5, 7]ĄĨ ĢīĄËĨØĨęĨŠĨÐĨŊĨÆĨęĨĒĄĘHeliobacteriaĄËĄ§ÎÐŋ§ÎēēŦšŲķÝÆąÍÍĄĪŧĀÁĮĪËđâĪĪīķžõĀĪōŧýĪÄļũđįĀŪšŲķÝĪĮĪĒĪęĄĪžįĪËÅÚūíÃæĪĮļŦĪÄĪŦĪëĄĨļũđįĀŪšŲķÝĪÎÃæĪĮÍĢ°ėĨđĨÝĨĒĄĘËĶŧŌĄËĪηÁĀŪĮ―ĪōÍĪ·ĪÆĪĪĪëĨ°Ĩ륞ĨŨĪĮĪĒĪęĄĪĪ―ĪÎĨđĨÝĨĒĪÏīĨÁįĪäĮŪĪËķŊĪĪÂŅĀĪōžĻĪđĄĨžįĪĘļũđįĀŪŋ§ÁĮĪÏBchl gĪČļÆĪÐĪėĪëĨŊĨíĨíĨÕĨĢĨëaĪËŧũĪŋÆČÆÃĪĘŋ§ÁĮĪĮĪĒĪëĄĨ·ÏÅýÅŠĪËĪÏĨ°ĨéĨāÍÛĀšŲķÝĪĮĪĒĪëChlostridiumĪĘĪÉĪŽÂ°ĪđĪëFirmicutesĖįĪËĘŽÎāĪĩĪėĪë[8, 9]ĄĨ ĪģĪÎĪčĪĶĪËČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏ·ÏÅýģØÅŠĪËĖįĪČĪĪĪĶÂįĪĪĘĘŽÎāģŽĩéĨėĨŲĨëĪĮđĪŊŧķšßĪ·ĪÆĪĪĪëĪÐĪŦĪęĪĮĪĘĪŊĄĪÂįĪĪĘĀļÍýģØÅŠĄĶ·ÁÂÖģØÅŠÂŋÍÍĀĪōŧýĪÃĪÆĪĪĪëĄĨĪģĪėĪÏŧĀÁĮČŊĀļ·ŋļũđįĀŪĪČĪĪĪĶĩĄĮ―ĪŽĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎÃæĪËĪÎĪßļŦ―ÐĪĩĪėĪëĪÎĪČÂÐūČÅŠĪĮĪĒĪëĄĨĪģĪÎĪčĪĶĪĘ·ÏÅýŋĘē―ģØÅŠÃÎļŦĪŦĪéĄĪŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎ―Ðļ―ĪËĀčÎĐĪÃĪÆČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪŽ―Ðļ―Ī·ĪÆĪŠĪęĄĪģîĪÄšŲķÝĪËĪŠĪąĪëČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎģÍÆĀĪÏĀļĘŠŧËĪÎÃæĪĮĪâķËĪáĪÆļÅĪĪĨĪĨŲĨóĨČĪĮĪĒĪëĪģĪČĪŽžĻšķĪĩĪėĪëĄĨ
ĢģĄĨŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎĨáĨŦĨËĨšĨā
ŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÏžįĪË°ĘēžĪÎĢģĪÄĪÎËėīÓÄĖ·ŋĨŋĨóĨŅĨŊžÁĪËĪčĪęđÔĪïĪėĪÆĪĪĪëĄĨĪ―ĪėĪÏĄĪļũē―ģØ·ÏII (PS II)ĄĪ Cytochrome b6f complex (Cyt b6f) Ī―Ī·ĪÆļũē―ģØ·ÏI (PS I)ĪĮĪĒĪë ĄĘFig. 1 ŧēūČĄË[10, 11]ĄĨ PS IIĪÏĨŊĨíĨíĨÕĨĢĨëĪōīÞĪāļũķîÆ°·ŋĪÎđÚÁĮĪĮĪĒĪęĄĪļũĪÎĨĻĨÍĨëĨŪĄžĪōÍøÍŅĪ·ŋåĪŦĪéÅÅŧŌĪōÃĨĪĪĨŨĨéĨđĨČĨĨÎĨóĄĘPQĄËĪČĪĪĪĶŧéÍÏĀĪÎÅÅŧŌąŋČÂÂÎĪËÅÅŧŌĪōÅÏĪđĄĨĪģĪÎĪČĪÅÅŧŌĪōÃĨĪïĪėĪŋŋåĪÏĨŨĨíĨČĨóĪČŧĀÁĮļķŧŌĪËĘŽĪŦĪėĪëĄĨĢēĘŽŧŌĪÎŋåĪŽĢīÅÅŧŌŧĀē―ĪĩĪėĪëĪģĪČĪËĪčĪęĢąĘŽŧŌĪÎĘŽŧŌūõŧĀÁĮĪŽČŊĀļĪđĪëĄĨPS IIÆâĪÎPQĪÏÅÅŧŌĪōžõĪąžčĪÃĪŋŧöĪËĪčĪęŋåÁĮĨĪĨŠĨóĄĘĨŨĨíĨČĨóĄËĪōËėģ°ŋåÁęĪčĪęĘáģÍĪ·ĄĪËėÃæĪō°ÜÆ°Ī·ĪÆĄĪCyt b6fĪËĪŋĪÉĪęÃåĪŊĄĨPQĪÏCyt b6fĪØÅÅŧŌĪōžõĪąÅÏĪđĪČĪČĪâĪËĄĪĨŨĨíĨČĨóĪōĄĘžõĪąžčĪÃĪŋĪÎĪČĪÏĩÕÂĶĪÎËėģ°ĪØĄËĨęĨ꥞ĨđĪđĪëĄĨCyt b6fĪŽžõĪąžčĪÃĪŋÅÅŧŌĪÏĨŨĨéĨđĨČĨ·ĨĒĨËĨó(PC)ĪČĪĪĪĶŋåÍÏĀĪÎÅÅŧŌąŋČÂÂÎĪËžęÅÏĪĩĪėĄĪPS IĪØąŋĪÐĪėĪëĄĨPS IĪÏPS IIÆąÍÍĨŊĨíĨíĨÕĨĢĨëĪōīÞĪāļũķîÆ°·ŋĪÎđÚÁĮĪĮĪĒĪęĄĪļũĨĻĨÍĨëĨŪĄžĪōÍøÍŅĪ·ĪÆPCĪŦĪéžõĪąžčĪÃĪŋÅÅŧŌĪōÅÅŧŌĘäÂÎÏĪΞåĪĪĨÕĨ§ĨėĨÉĨĨ·ĨóĄĘFdĄËĪËÅÏĪđĄĨĪ―Ī·ĪÆīÔļĩĪĩĪėĪŋFdĪŽNADP+ĪËÅÅŧŌĪōÍŋĪĻĄĪNADPHĪŽšîĪéĪėĪëĄĨĀļĀŪĪĩĪėĪŋNADPHĪÏīÔļĩÎÏĪČĪ·ĪÆÚŧĀļĮÄęĪäĘŠžÁĀļŧšĪËÍŅĪĪĪéĪėĪëĄĨĪÞĪŋĄĪĪģĪėĪéËėĨŋĨóĨŅĨŊžÁīÖĪōÅÅŧŌĪŽ°ÜÆ°ĪđĪëīÖĪËËėĪōģÖĪÆĪŋŋåÁĮĨĪĨŠĨóĄĘĨŨĨíĨČĨóĄËĪÎļûĮÛĪŽ·ÁĀŪĪĩĪėĄĪËėīÓÄĖ·ŋĪÎATPđįĀŪđÚÁĮĪŽĪģĪÎļûĮÛĪōÍøÍŅĪ·ATPĪÎĀļŧšĪōđÔĪĶĄĨĪģĪÎĪčĪĶĪĘīÔļĩÎÏĄĘNADPHĄËĪČĨĻĨÍĨëĨŪĄž(ATP)ĪÎĀļŧšĪōđÔĪĶĪŋĪáĪËËėĨŋĨóĨŅĨŊžÁĪŽÅÅŧŌĪΞõĪąÅÏĪ·ĪōĪđĪëĨ·ĨđĨÆĨāĪōĄÖÅÅŧŌÅÁÃĢ·ÏĄŨĪČļÆĪÖĪŽĄĪŧĀē―īÔļĩĨÝĨÆĨóĨ·ĨãĨëĪō―ÄžīĪČĪ·ĪÆ ĪģĪÎÅÅŧŌĪÎÎŪĪėĪōÉÁĪĪĪŋĪâĪÎĪŽFig. 2ĪĮĪĒĪëĄĨÅÅŧŌĪÏŧĀē―īÔļĩĨÝĨÆĨóĨ·ĨãĨëĪÎĨÞĨĪĨĘĨđÂĶĄĘŋÞĪÎūåĘýĄËĪŦĪéĨŨĨéĨđÂĶĄĘēžĘýĄËĪØĪČžøžõĪĩĪėĪëĪŽĄĪĪ―ĪÎĩÕĘýļþĪØĪΰÜÆ°ĪËĪÏĨĻĨÍĨëĨŪĄžĪŽÉŽÍŨĪČĪĩĪėĪëĄĨŧĀÁĮČŊĀļ·ŋļũđįĀŪĪËĪŠĪĪĪÆĪÏÅÅŧŌĘäÂÎÏĪÎķŊĪĪŋå(+820 mV)ĪŦĪéÅÅŧŌĘäÂÎÏĪΞåĪĪNADP(-320 mV)ĪØÅÅŧŌĪŽžõĪąÅÏĪĩĪėĪëīÖĪËļũĪËĪčĪëÅÅŧŌĪÎĨÝĨóĨŨĨĒĨÃĨŨĪŽÆóÅŲđÔĪïĪėĪÆĪĪĪëĄĨĪģĪÎÆóÅŲĪÎĨÝĨóĨŨĨĒĨÃĨŨĪÎĪŋĪáĪËĢēžïÎāĪÎļũē―ģØ·ÏĄĘPS IIĪČPS IĄËĪŽīØÍŋĪ·ĪÆĪĪĪëĪÎĪĮĪĒĪëĄĨ
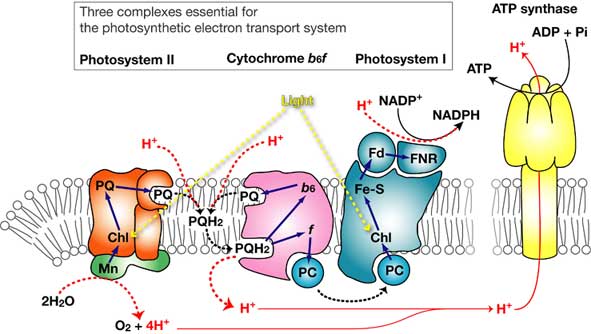
ĢīĄĨČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎļũē―ģØ·Ï
ŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪŽPS IIĪČPS IĪČĪĪĪĶĢēžïÎāĪÎļũē―ģØ·ÏĪōŧýĪÃĪÆĪĪĪëĪÎĪËÂÐĪ·ĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏĪ―ĪÎĪĶĪÁĪÉĪÁĪéĪŦ°ėĘýĪÎļũē―ģØ·ÏĪ·ĪŦŧýĪŋĪĘĪĪ(Fig. 2 ŧēūČ) [12]ĄĨÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪČđČŋ§ļũđįĀŪšŲķÝĪÏPS IIĪËÎāŧũĪ·Īŋļũē―ģØ·ÏĪÎĪßĪĮļũđįĀŪĪōđÔĪÃĪÆĪŠĪęĄĪÎÐŋ§ÎēēŦšŲķÝĪČĨØĨęĨŠĨÐĨŊĨÆĨęĨĒĪÏPS IÎāŧũĪÎļũē―ģØ·ÏĪĀĪąĪōŧýĪÄĄĨļũđįĀŪĪËĪŠĪĪĪÆĢēĪÄĪÎļũē―ģØ·ÏĪōđįĪïĪŧŧýĪÄÉŽÍŨĪÏĪĘĪŊĄĪĪÉĪÁĪéĪŦ°ėĘýĪĀĪąĪĮĪâĨĻĨÍĨëĨŪĄž(ATP)ĪČīÔļĩÎÏ(ČóŧĀÁĮČŊĀļ·ŋĪËĪŠĪĪĪÆĪÏNADH)ĪÎĀļŧšĪŽ――ĘŽĪËēÄĮ―ĪĘĪÎĪĮĪĒĪëĄĨÂįĪĪĘ°ãĪĪĪÏÅÅŧŌÅÁÃĢĪÎÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆŋåĪōŧČĪĶĪģĪČĪŽ―ÐÍčĪĘĪĪĪČļĀĪĶĪģĪČĪĮĪĒĪëĄĨĪ―ĪėĪŽļũđįĀŪĪËĪŠĪĪĪÆŧĀÁĮĪÎČŊĀļĪōČžĪïĪĘĪĪĪģĪČĪÎÍýÍģĪĮĪĒĪëĪŽĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆŋåĪĮĪÏĪĘĪŊĄĪÎēē―ŋåÁĮÅųĪÎīÔļĩÅŠÎēēŦē―đįĘŠĄĪŋåÁĮĄĪÍĩĄĘŠĪĘĪÉĪōÍŅĪĪĪëĄĨŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪĮĪĒĪëĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎPS IIĪÏŋåĪŦĪéÅÅŧŌĪō°úĪČīĪŊĪŋĪáĪËÆÃĘĖĪĘđ―ÂĪÂÎĪōČŊÃĢĪĩĪŧĪÆĪĪĪë[13]. Ī―ĪėĪÏĢīĪÄĪÎĨÞĨóĨŽĨóļķŧŌĪōīÞĪāĨæĨËĨÃĨČĪĮĄĪĢīÅÅŧŌŧĀē―ĪōžõĪąĪÆĪĪĪëīÖĪÎÉÔī°ÁīĪĘŧĀē―ūõÂÖĪËĪŠĪŦĪėĪÆĪĪĪëŋåĪō°ÂÄęē―ĪĩĪŧĪëĪŋĪáĪÎđ―ÂĪĪĮĪĒĪëĪČĪĩĪėĪÆĪĪĪëĄĨÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪČđČŋ§ļũđįĀŪšŲķÝĪÏPS IIĪËķËĪáĪÆÎāŧũĪ·Īŋļũē―ģØ·ÏĪōŧýĪÄĪŽĄĪŧĀÁĮČŊĀļĪËīØÍŋĪđĪëĪģĪÎĨæĨËĨÃĨČĪō·įĪŊĪŋĪáĄĪŋåĪōÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆÍøÍŅ―ÐÍčĪĘĪĪ[14, 15]ĄĨÂūĘýĄĪPS IĪÏPS IIĪČČæĪŲĪÆÄãĪĪŧĀē―īÔļĩÅÅ°ĖĪËÅŽąþĪ·Īŋļũē―ģØ·ÏĪĮĪĒĪęĄĪÅÅ°ĖĪÎÄãĪĪÅīÎēēŦĨŊĨéĨđĨŋĄžĄĘFe-SĄËĪōīÞĪßFdĪō·ÐĪÆNADĪōÄūĀÜīÔļĩ―ÐÍčĪëĪČĪĪĪĶÆÃħĪōŧýĪÄĪâĪÎĪĮĪĒĪë[16]ĄĨĪģĪÎPS IĪËÎāŧũĪ·Īŋļũē―ģØ·ÏĪÎĪßĪĮļũđįĀŪĪōđÔĪĶÎÐŋ§ÎēēŦšŲķÝĪäĨØĨęĨŠĨÐĨŊĨÆĨęĨĒĪÏĄĪĪ―ĪÎļũē―ģØ·ÏžŦÂÎĪÎŧĀē―īÔļĩÅÅ°ĖĪŽÄãĪđĪŪĪÆÅÅŧŌĘäÂÎÏĪÎķŊĪĪŋåĪŦĪéÅÅŧŌĪō°úĪČīĪŊĪģĪČĪÏÉÔēÄĮ―ĪĮĄĪŋåĪōÅÅŧŌķĄÍŋÂÎĪČĪđĪëĪģĪČĪÏ―ÐÍčĪĘĪĪĄĨ
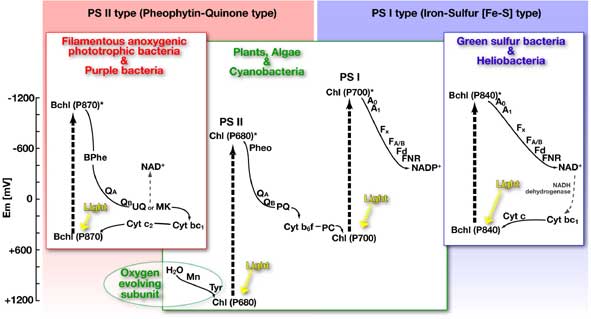
ĢĩĄĨļũē―ģØ·ÏĪÎŋĘē―
ļũđįĀŪĪÎĢē·ŋž°ĄĘŧĀÁĮČŊĀļ·ŋĪČČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĄËĪÎĖĀģÎĪĘ°ãĪĪĪΰėĪÄĪÏĄĪŧĀÁĮČŊĀļ·ŋļũđįĀŪĪĮĪÏÆóĪÄĪÎļũē―ģØ·ÏĄĘPS IIĪČPS IĄËĪōÍŅĪĪĪëĪÎĪËÂÐĪ·ĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪĮĪÏĪ―ĪÎĪÉĪÁĪéĪŦ°ėĘýĪ·ĪŦÍŅĪĪĪĘĪĪĪČĪĪĪĶĪģĪČĪĮĪĒĪëĄĨĪÄĪÞĪęĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪŽŧĀÁĮČŊĀļĪČĪĪĪĶÆÃĘĖĪĘĩĄđ―ĪōģÍÆĀĪđĪëĪŋĪáĪËĪÏĄĪ°ėĘýĪĀĪąĪĮĪâļũđįĀŪÅŠĪË――ĘŽĪËĀļ°éēÄĮ―ĪĘÆóĪÄĪÎļũē―ģØ·ÏĪōđįĪïĪŧŧýĪÄÉŽÍŨĪŽĪĒĪÃĪŋĪÎĪĀĪíĪĶĪČđÍĪĻĪÆĪĪĪëĄĨÆóĪÄĪÎļũē―ģØ·ÏĪÏļũĨĻĨÍĨëĨŪĄžĪōÍøÍŅĪ·ÅÅŧŌĘäÂÎÏĪÎķŊĪĪĪâĪÎĪŦĪéžåĪĪĪâĪÎĪËÅÅŧŌĪōžõĪąÅÏĪđĪČĪĪĪÃĪŋĩĄĮ―ĪŦĪéļŦĪėĪÐÆąžÁĪĮĪĒĪëĪČļĀĪĻĪëĪŽĄĪĪ―ĪėĪéĪÎŧĀē―īÔļĩÅÅ°ĖÂÓĪÏ°ÛĪĘĪÃĪÆĪŠĪęĄĪĨĨÎĨóĪÎīÔļĩĪōđÔĪĶPS IIĪŽŧĀē―ÅŠĄĘŧĀē―īÔļĩÅÅ°ĖÂÓĪŽđâĪáĄËĪĘĪÎĪËÂÐĪ·ĄĪFdĪōīÔļĩĪđĪëPS IĪÏīÔļĩÅŠĄĘŧĀē―īÔļĩÅÅ°ĖÂÓĪŽÄãĪáĄËĪĘ·đļþĪōŧýĪÃĪÆĪĪĪëĄĨĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪËļŦĪéĪėĪëPS IIĪÏĄĪČóŧĀÁĮČŊĀļ·ŋšŲķÝĪÎŧýĪÄÎāŧũļũē―ģØ·ÏĪËČæĪŲÅÅŧŌĘäÂÎÏĪŽķŊĪŊĪĘĪÃĪÆĪŠĪęĄĪđđĪËĪÞĪŋĄĪĨÞĨóĨŽĨóļķŧŌĪōīÞĪāĄÖŧĀÁĮČŊĀļĨæĨËĨÃĨČĄŨĪÏŋåĪŦĪéÅÅŧŌĪō°úĪČīĪŊĪģĪČĪŽ―ÐÍčĪëĪÞĪĮĪËĪĘĪÃĪÆĪĪĪëĄĨĪģĪÎĪčĪĶĪĘPS IIĪÎÅÅŧŌžõÍÆÂĶĪÎđâŧĀē―īÔļĩÅÅ°ĖĘýļþĪØĪÎĨ·ĨÕĨČĪÏĄĪĪâĪĶ°ėĪÄĪÎļũē―ģØ·ÏĪōđįĪïĪŧŧýĪÃĪÆĪĪĪëĪČĪĪĪĶĄÖĨĻĨÍĨëĨŪĄžÅŠĪĘĪæĪČĪęĄŨĪŽĀļĪß―ÐĪ·ĪŋĄĘČŊÅļÅŠĪĘĄËĘŅē―ĪĮĪĒĪëĪČļĀĪĻĪëĪŦĪâÃÎĪėĪĘĪĪĄĨFig. 3ĪÏ16S rRNAĮÛÎóĪËīðĪÅĪĪĪŋģÆļũđįĀŪšŲķÝĪηÏÅýŋĘē―ÅŠīØ·ļĪōÉ―Ī·Īŋ·ÏÅýžųĪĮĪĒĪëĄĨŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÏĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪηÏÅýĪËĪ·ĪŦļŦĪéĪėĪĘĪĪĪģĪČĪŦĪéĄĪĪģĪÎĨ°Ĩ륞ĨŨĪŽĘŽīôĪ·Īŋ°ĘđßĪËģÍÆĀĪĩĪėĪŋĪâĪÎĪĮĪĒĪëĪČđÍĪĻĪëĪģĪČĪŽĪĮĪĪëĄĨPS IĨŋĨĪĨŨĪÎļũē―ģØ·ÏĪÎĪßĪōŧýĪÄÎÐŋ§ÎēēŦšŲķÝĪČĨØĨęĨŠĨÐĨŊĨÆĨęĨĒĪÏ·ÏÅýžųūåĪĮĪÏÎĨĪėĪÆÂļšßĪ·ĪÆĪĪĪëĄĨĪģĪÎĪģĪČĪÏPS IĨŋĨĪĨŨĪÎļũē―ģØ·ÏĪŽĄĪĪģĪėĪéÎū·ÏÅýĪŽĘŽīôĪđĪë°ĘÁ°ĪËģÍÆĀĪĩĪėĪÆĪĪĪŋĪģĪČĪōžĻšķĪđĪëĄĨĪÞĪŋĄĪPS IIĨŋĨĪĨŨĪÎļũē―ģØ·ÏĪÎĪßĪōŧýĪÄÁĄ°ÝūõČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪČđČŋ§šŲķÝĪÏ·ÏÅýÅŠĪËđđĪËģÖĪŋĪÃĪÆĪŠĪęĄĪĪģĪÎĨŋĨĪĨŨĪÎļũē―ģØ·ÏĄĘPS IIĄËĪÎĩŊļķĪŽPS IĪčĪęļÅĪĪĪģĪČĪōžĻšķĪđĪëĄĨ
ĪģĪÎĪčĪĶĪË16S rRNAĮÛÎóĪËīðĪÅĪŊēōĀÏĪĮĪÏĄĪŧĀÁĮČŊĀļ·ŋļũđįĀŪĪËĀčĪóĪļĪÆČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪŽ―Ðļ―Ī·ĄĪĪÞĪŋļũē―ģØ·ÏĪÎģÍÆĀ―į―øĪÏPS II ĒÍ PS IĪÎ―įĪĮĪĒĪÃĪŋĪģĪČĪŽžĻĪĩĪėĪëĄĨÂūĘýĄĪļũē―ģØ·ÏĪËīÞĪÞĪėĪëŋ§ÁĮĄĘBchlĄËĪÎđįĀŪđÚÁĮĪÎĨĒĨßĨÎŧĀĮÛÎóĪÎČæģÓēōĀÏ[17, 18]ĪäļũđįĀŪšŲķÝĪÎÁīĨēĨÎĨāĮÛÎóĪōÂÐūÝĪČĪ·Īŋ·ÏÅýŋĘē―ÅŠļĶĩæ[19]ĪŦĪéĪÏĄĪĪģĪģĪĮžĻĪ·Īŋ16S rRNAĮÛÎóĪŦĪéĪÎēōĀÏ·ëēĖĪČ°ÛĪĘĪëŋĘē―·ÐÏĐĪŽžĻšķĪĩĪėĪëĪģĪČĪâÂŋĪĪĄĨĪ·ĪŦĪ·ĄĪĪÉĪÎÍÍĪĘēōĀÏĪËĪŠĪĪĪÆĪâĄÖČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÏŧĀÁĮČŊĀļ·ŋļũđįĀŪĪËĀčđÔĪ·ĪÆģÍÆĀĪĩĪėĪÆĪĪĪëĪģĪČĄŨ ĪČĄĪĄÖPS IIĪÎĩŊļķĪÏPS IĪÎĪ―ĪėĪčĪęĪâļÅĪĪĄŨĪČĪĪĪĶ·ëēĖĪÎÆĀĪéĪėĪëĪģĪČĪŽÂŋĪŊĄĪĪ―ĪėĪéĪηëÏĀĪÏÂÅÅöĪĘĪâĪÎĪĮĪĒĪëĪČļĀĪÃĪÆĪčĪĪĪČđÍĪĻĪÆĪĪĪëĄĨ
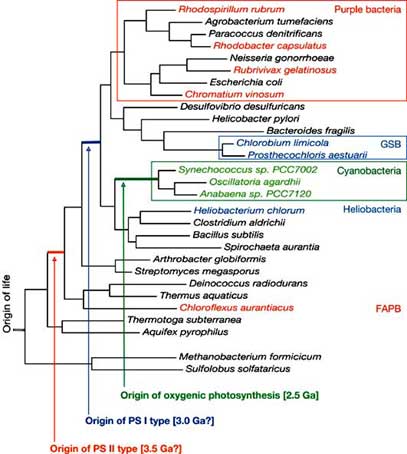
ĢķĄĨļũđįĀŪĪÎŋĘē―ēūĀâ
ĪģĪÎĪčĪĶĪĘēōĀÏ·ëēĖĪōƧĪÞĪĻĪÆĄĪļũđįĀŪĄĘļũē―ģØ·ÏĄËĪÎŋĘē―ēáÄøĪōŋäÄęĪ·ĪŋĪâĪÎĪŽFig. 4ĪËžĻĪ·ĪŋŋĘē―ēūĀâĪĮĪĒĪ륧ĢąĄË―éīüĀļĖŋÂÎĪËĪŠĪąĪëļũđįĀŪĩĄĮ―ĪÎģÍÆĀ
ĀļĖŋĪÎÎōŧËĪÎ―éīüĪÎÃĘģŽĪĮļũđįĀŪĩĄĮ―ĪŽģŦČŊĪĩĪėĪŋĪĮĪĒĪíĪĶĪŽĄĪĪ―ĪÎļũđįĀŪĪÏPS IIĨŋĨĪĨŨĪÎļũē―ģØ·ÏĪōģËĪČĪ·ĪŋĪâĪÎĪĮĪĒĪëĄĨĪģĪÎPS IIĨŋĨĪĨŨĪÏĨĨÎĨóĪōīÞĪāŧĀē―īÔļĩÅÅ°ĖĪÎĪĒĪÞĪęÄãĪŊĪĘĪĪļũē―ģØ·ÏĪĮĪĒĪęĄĪNADĪōÄūĀÜīÔļĩ―ÐÍčĪĘĪĪĪģĪČĪŦĪéĄĪĄÖĨĻĨÍĨëĨŪĄžĀļŧšĩĄĮ―ĪČĪ·ĪÆ――ĘŽĪĮĪĒĪëĪŽĄĪĀöÎýĪĩĪėĪÆĪĪĪĘĪĪļũđįĀŪīïīąĄŨĪĮĪĒĪëĪČļĀĪĻĪëĄĨ
ĢēĄËīÄķĪËĪčĪęÅŽąþĪ·Īŋļũē―ģØ·ÏĪÎģŦČŊ
ĪäĪŽĪÆĄĪPS IĨŋĨĪĨŨĪÎļũē―ģØ·ÏĪŽģŦČŊĪĩĪėĪëĄĨĪģĪÎPS IĨŋĨĪĨŨĪÏFdĪō·ÐĪÆNADĪōÄūĀÜīÔļĩ―ÐÍčĪëÄãŧĀē―īÔļĩÅÅ°Ė·ŋĪĮĪĒĪëĄĨĪģĪÎĨŋĨĪĨŨĪÎļũđįĀŪÁõÃÖĪōŧýĪÃĪŋĀļĘŠĪÏĄĪŧĀÁĮĪÎÂļšßĪ·ĪĘĪĪ·ųĩĪÅŠļÅÃÏĩåīÄķĪËĪŠĪĪĪÆĄĪĀļÂÎĘŠžÁĀļŧšĪÎĪŋĪáĪÎīÔļĩÎÏĪÎĀļĀŪĪČÚŧĀļĮÄęĪōļúÎĻĪÎĪčĪŊĄĘÅÅŧŌÅÁÃĢĪŽÂÚĪëŧöĪĘĪŊĄËđÔĪĶĪģĪČĪŽ―ÐÍčĪëĄĨĪģĪÎĪčĪĶĪĘĪģĪČĪŦĪéĄĪĀčĪËļ―ĪėĪŋPS IIĨŋĨĪĨŨĪÎļũđįĀŪšŲķÝĪËÂÐĪ·ĄĪĪ―ĪÎÅöŧþĄĪ°ĩÅÝÅŠĪËÍĨĀŠĪËĪĘĪÃĪŋĪâĪÎĪČÁÛÁü―ÐÍčĪëĄĨ
ĢģĄËŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎ―Ðļ―
ļÅÃÏĩåĪËĪŠĪĪĪÆĪÏŧĀÁĮČŊĀļĪōČžĪïĪĘĪĪļũđįĀŪĪōđÔĪĶšŲķÝĪÎŧþÂåĪŽĪŦĪĘĪęÄđĪĪËÅÏĪęÂģĪĪĪŋĪâĪÎĪČđÍĪĻĪéĪėĪëĄĨĪ―Ī·ĪÆĄĪšĢĪčĪęĢēĢĩēŊĮŊĪÛĪÉÁ°ĪË―éĪáĪÆÃÏĩåūåĪËŧĀÁĮČŊĀļ·ŋļũđįĀŪĪŽÅÐūėĪđĪëĄĨĪđĪĘĪïĪÁĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎ―Ðļ―ĪĮĪĒĪëĄĨĪÞĪšĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎÁÄĀčžïĪÎÃæĪĮÆóžïÎāĪÎļũē―ģØ·ÏĄĘPS IIĪČPS IĄËĪŽĪŌĪČĪÄĪΚŲËĶÃæĪËķĶÂļĪđĪëĪČĪĪĪÃĪŋĨĪĨŲĨóĨČĪŽĩŊĪģĪÃĪŋĪČđÍĪĻĪéĪėĪëĄĨĪģĪÎĨÏĨĪĨÖĨęĨÃĨÉē―ĪÏĄĪļũđįĀŪ°äÅÁŧŌĪÎÅÁČÂĪËĪčĪëĪâĪÎĪČđÍĪĻĪéĪėĪëĄĨĪâĪČĪâĪČPS IIĨŋĨĪĨŨĪÎļũđįĀŪšŲķÝĪËPS IĪΰäÅÁŧŌĪŽÅÁČÂĪ·ĪŋĪŦĄĪĪ―ĪÎĩÕĪĮĪĒĪëĪÎĪŦĪÏĘŽĪŦĪéĪĘĪĪĪŽĄĪ·ëēĖĪČĪ·ĪÆĪŌĪČĪÄĪΚŲËĶĪÎÃæĪËĢēžïÎāĪÎļũđįĀŪ·ÏĪŽšŪšßĪđĪëĪģĪČĪËĪĘĪÃĪŋĄĘĪģĪÎŧþĪΰäÅÁŧŌÅÁČÂĪÎĨáĨŦĨËĨšĨāĪËĪÄĪĪĪÆÁÛÁüĪđĪëĪčĪęÂūĪËĪĘĪĪĪŽĄĪļũđįĀŪšŲķÝĪŽÂūžïÎāĪÎļũđįĀŪšŲķÝĪōšŲËĶĪīĪČžčĪęđþĪóĪĀŧöĄĪĪđĪĘĪïĪÁPhagocytosis ĪËÞĪōČŊĪ·ĪŋĪÎĪĮĪÏĪĘĪĪĪŦĪČÉŪžÔĪÏđÍĪĻĪÆĪĪĪëĄĨļ―ÂļĪđĪëšŲķÝĪĮĪÏĪģĪÎĪčĪĶĪĘPhagocytosisĪÏĪÛĪČĪóĪÉļŦĪéĪėĪĘĪĪĪŽĄĪļũđįĀŪĪōđÔĪĶŋŋģËĀļĘŠĪĮĪĒĪëÁôÎāĪËĪŠĪĪĪÆĪÏÁôÎāÆąŧÎĪÎĄÖŋĐĪĪđįĪĪĄŨĪŽŋôÂŋĪŊĘóđðĪĩĪėĪÆĪĪĪëĄĨÅöÁģĄĪŋŋģËĀļĘŠĪČļķģËĀļĘŠĪōÆąÎóĪËĩÄÏĀĪđĪëĪģĪČĪÏ―ÐÍčĪĘĪĪĪģĪČĪÏūĩÃÎĪÎūåĪĮĪÏĪĒĪëĪŽĄĶĄĶĄĶĄËĄĨšĮ―éĄĪĪģĪėĪéÆóĪÄĪÎļũē―ģØ·ÏĪÏĪŌĪČĪÄĪΚŲËĶÆâĪĮÆČÎĐĪËÆŊĪĪĪÆĪĪĪŋĪŽĄĪĪ―ĪÎĨĻĨÍĨëĨŪĄžÅŠĪĘÍūÍĩĪŦĪéĪŦĄĪPS IIĪÎŋĘē―ĪŽÂįĪĪŊŋĘĪāĪģĪČĪËĪĘĪëĄĨĪ―Ī·ĪÆĄĪĢīĪÄĪÎĨÞĨóĨŽĨóļķŧŌĪōīÞĪāŧĀÁĮČŊĀļĨæĨËĨÃĨČĪÎģÍÆĀĪČĪĪĪĶÂčĢēĪÎĨĪĨŲĨóĨČĪŽĩŊĪģĪëĪÎĪĮĪĒĪëĄĨĪđĪĘĪïĪÁĄĪĪģĪėĪģĪ―ĪŽŧĀÁĮČŊĀļ·ŋļũđįĀŪĩĄđ―ĪÎī°ĀŪĪĮĪĒĪęĄĪŋåĪōÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆŧČÍŅĪđĪëĪģĪČĪÎ―ÐÍčĪëĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎĀčÁÄ·ŋĪŽ―Ðļ―ĪđĪëĪģĪČĪČĪĘĪÃĪŋĄĨÂūĘýĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏĄĪÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆÎēē―ŋåÁĮĪĘĪÉēÐŧģĀĪÎīÔļĩĘŠžÁĪ·ĪŦÍøÍŅ―ÐÍčĪĘĪĪĪŋĪáĄĪĪģĪėĪŽĀļ°éīÄķĪōļÂÄęĪđĪëĪģĪČĪČ·ŌĪŽĪëēÄĮ―ĀĪÏÂįĪĪĪĄĨĪ·ĪŦĪ·ĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÏĪÉĪģĪËĪĮĪâÂļšßĪđĪëŋåĪōÅÅŧŌķĄÍŋÂÎĪČĪ·ĪÆÍŅĪĪĪëĪģĪČĪŽ―ÐÍčĪëĪŋĪáĄĪÃÏĩåūåĪÎÍÍĄđĪĘīÄķÃæĪĮĀļ°éĪ·ÆĀĪëĪčĪĶĪËĪĘĪÃĪŋĄĨŋåĪËēÃĪĻĄĪÚŧĀĨŽĨđĪČļũĪŽĪĒĪėĪÐīðËÜÅŠĪËÁýŋĢēÄĮ―ĪĮĪĒĪëĪŋĪáĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÏĄĪÃÏĩåÉ―ÁØÉôĪÎĪÛĪÜÁīĪÆĪÎŋå·ũĪËĄĪĪ―ĪÎĀļ°éĪËĪČĪÃĪÆĪÎÉŽÍŨ――ĘŽūō·ïĪōļŦ―ÐĪŧĪŋĪËĪÁĪŽĪĪĪĘĪĪĄĨĪģĪĶĪ·ĪÆĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÎÂįČËŋĢĪŽÃÏĩåīÄķĪōĘŅĪĻĪëĪÞĪĮĪËđĪŽĪÃĪÆĪĪĪŊĪģĪČĪČĪĘĪÃĪŋĄĨ
ĢīĄËĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪŽČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪËĩÚĪÜĪ·ĪŋąÆķÁ
Ĩ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪÏŧĀÁĮČŊĀļ·ŋļũđįĀŪĪČĪĪĪĶķËĪáĪƷКŅÅŠĪĮļúÎĻÅŠĪĘĨĻĨÍĨëĨŪĄžģÍÆĀžęÃĘĪōÍŅĪĪĪÆĄĪĪ―ĪÎŋôĪōÁýĪäĪ·ĪÆĪĪĪÃĪŋĄĨĪ―ĪÎČŋĖĖĄĪĮŅīþĘŠĪČĪ·ĪÆÂįÎĖĪÎŧĀÁĮĪŽĮÓ―ÐĪĩĪėĪëĪŋĪáĄĪĪäĪŽĪÆÃÏĩåĩŽĖÏĪĮĪÎīÄķĘŅē―ĪŽ°úĪĩŊĪģĪĩĪėĪëĪģĪČĪČĪĘĪëĄĨĮÓ―ÐĪĩĪėĪŋŧĀÁĮĪÏĄĪÅö―éĄĪģĪŋåÃæĪËÂŋÎĖĪËīÞĪÞĪėĪÆĪĪĪëīÔļĩÅīĪĘĪÉĪÎīÔļĩĘŠžÁĪÎŧĀē―ĪČĪĪĪĶ·ÁĪĮ―čÍýĪĩĪėĪÆĪĪĪŋĪŽĄĪĪäĪŽĪÆĪ―ĪÎĨęĨķĄžĨÐĄžĪâÄėĪōŋÔĪĄĪÂįĩĪÃæĪË°îĪėĪÆĪĪĪŊĪģĪČĪČĪĘĪëĄĨĪ―Ī·ĪÆĄĪšĢĪčĪęĢēĢēēŊĮŊĪŦĪéĢąĢđēŊĮŊÁ°ĪËÂįĩĪÃæĪÎŧĀÁĮĘŽ°ĩĪŽÂįĪĪŊūåūšĪ·ĄĪļ―šßĪÎĢąĢĩĄóĪËÃĢĪđĪëĪÛĪÉĪËĪĘĪÃĪŋ[20]ĄĨÃÏĩåīÄķĪÏĪÉĪóĪÉĪóŧĀē―ÅŠĪËĪĘĪÃĪÆĪĪĪĄĪ·ųĩĪÅŠīÄķĪĮŋĘē―Ī·ĪÆĪĪŋĀļĘŠÃĢĪđĪŲĪÆĪËŋžđïĪĘ°ąÆķÁĪōĩÚĪÜĪđĪģĪČĪČĪĘĪÃĪŋĄĨ·ųĩĪūō·ïēžĪĮĀļĪĪëĪâĪÎĪËĪČĪÃĪÆŧĀÁĮĪÏķŊÎõĪĘŧĀē―šÞĄĪĪđĪĘĪïĪÁÆĮĄĪĪËēáĪŪĪĘĪĪĪŦĪéĪĀĄĨĪģĪÎČïģēĪÏÅöÁģĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪËĩÚĪÖĄĨČāĪéĪÏĪßĪĘŧĀÁĮĪÎąÆķÁĪÎĩÚĪÐĪĘĪĪ·ųĩĪÅŠīÄķĪØĪČÄÉĪĪĪäĪéĪėĪëĪģĪČĪČĪĘĪÃĪŋĄĨĪ·ĪŦĪ·ĄĪPS IIĨŋĨĪĨŨĪÎļũđįĀŪšŲķÝĪÏĪ―ĪÎļũđįĀŪÁõÃÖĪŽPS IĨŋĨĪĨŨĪÎĪâĪÎĪËĪŊĪéĪŲŧĀē―ÅŠĘýļþĪËīóĪÃĪÆĪĪĪŋĪŋĪáĄĪĪģĪÎĪčĪĶĪĘŧĀē―ÅŠīÄķĪËĪĒĪëÄøÅŲÂŅĪĻĪëĪģĪČĪŽ―ÐÍčĪŋĄĨĪĩĪéĪËĪÏļũđįĀŪÅÅŧŌÅÁÃĢĪËīØĪïĪÃĪÆĪĪĪŋĪĪĪŊĪÄĪŦĪÎđÚÁĮĪōÎŪÍŅĪ·ĪÆĄĪšĮ―ŠÅŠĪËŧĀÁĮĪËÅÅŧŌĪōžÎĪÆĄĪĖĩÆĮĪĘŋåĪØĪČīÔļĩĪđĪëÅÅŧŌÅÁÃĢ·ÏĄĘĪđĪĘĪïĪÁŧĀÁĮļÆĩÛĄËĪōģÎÎĐĪđĪëĪģĪČĪËĀŪļųĪđĪëĪâĪÎĪâļ―ĪėĪŋĄĨĪģĪėĪËÂÐĪ·ĪÆPS IĨŋĨĪĨŨĪÎļũđįĀŪšŲķÝĪÏĄĪĪ―ĪÎļũđįĀŪÁõÃÖĪŽ·ųĩĪīÄķĪËī°ÁīĪËÅŽąþĪ·ĪÆĪŠĪęĄĪŧĀÁĮĪËÂÐĪđĪëķËĪáĪÆđâĪĪīķžõĀĪōŧýĪÄĪŋĪáĪËŧĀē―ÅŠīÄķĪĮĪÏĀļ°éĪđĪëĪģĪČĪÏÉÔēÄĮ―ĪĮĪĒĪÃĪŋĄĨĪčĪÃĪÆĄĪŧĀÁĮĪÎĪĒĪëīÄķĪË―įąþ―ÐÍčĪšĪËĄĪ·ųĩĪÅŠīÄķĪËÎąĪÞĪęÂģĪąļ―šßĪËŧęĪëĪģĪČĪČĪĘĪÃĪŋĄĨ
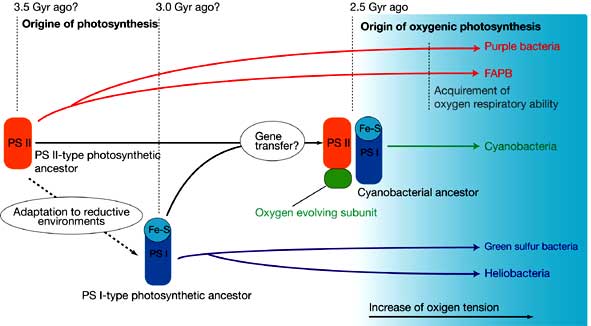
Ģ·ĄĨČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÎļÅÃÏĩåīÄķĪĮĪÎĖōģä
ūåĪËžĻĪ·ĪŋļũđįĀŪĪÎŋĘē―ēūĀâĪÏĄĪļũđįĀŪÁõÃÖĪÎĘŽŧŌđ―ÂĪĄĶĩĄđ―ĪÎČæģÓĪŠĪčĪÓĘŽŧŌŋĘē―ģØÅŠļĶĩæĨĮĄžĨŋĪËīðĪÅĪĄĪÉŪžÔĪŽÉÁĪŊČųĀļĘŠĀĪģĶĪËļŦĪéĪėĪëļũđįĀŪĩĄĮ―ģÍÆĀĪÎĄÖĨ·ĨĘĨęĨŠĄŨĪĮĪĒĪëĄĨĪģĪÎēūĀâĪΞÂūÚĪËīØĪ·ĪÆĪÏĄĪÅöÁģĄĪđđĪĘĪëĨĮĄžĨŋĪÎĀŅĪß―ÅĪÍĪČēōĀÏĪŽÉŽÍŨĪČĪĩĪėĪëĪĮĪĒĪíĪĶĪ·ĄĪĪÞĪŋŋ·ĪŋĪËÆĀĪéĪėĪŋŧöžÂĪŽĪģĪÎēūĀâĪōšŽÄėĪŦĪéĘĪĪđĪģĪČĪâĪĒĪęÆĀĪëĪĀĪíĪĶĄĨĪ·ĪŦĪ·ĄĪĄÖŧĀÁĮČŊĀļ·ŋļũđįĀŪĪÎĩŊļķĪŽČæģÓÅŠŋ·Ī·ĪŊĄĪĪ―ĪėĪËĀčđÔĪ·ĪÆČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĪŽģÍÆĀĪĩĪėĪÆĪĪĪŋĄŨĪČĪĪĪĶĪģĪČĪÏĄĪĩŋĪĪĪÎĪĘĪĪŧöžÂĪÎĪŌĪČĪÄĪČÉŪžÔĪÏđÍĪĻĪëĄĨĪÄĪÞĪęŧĀÁĮČŊĀļ·ŋļũđįĀŪĪŽ―Ðļ―ĪđĪë°ĘÁ°ĪÎÃÏĩåĪĮĪÏĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪŽēŋēŊĮŊĪâĪÎÄđĪĪËÅÏĪęĀļÂļĪ·ĪÆĄĘČËĖÐĪ·ĪÆĄËĪĪĪŋĪÎĪĮĪĒĪëĄĨĪĮĪÏĄĪļÅÃÏĩåĪÎČųĀļĘŠĀļÂÖ·ÏĪËĪŠĪĪĪÆĄĪĪģĪėĪéĪÎļũđįĀŪšŲķÝĪÏĪÉĪÎÍÍĪĘĖōģäĪōÃīĪÃĪÆĪĪĪŋĪÎĪĮĪĒĪíĪĶĪŦĄĨžþÃÎĪÎĪģĪČĪĮĪĒĪëĪŽĄĪĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒĪŽÅÐūėĪđĪë°ĘÁ°ĪÎÃÏĩåīÄķĪËĪÏĄĪĘŽŧŌūõŧĀÁĮĪÏĪÛĪČĪóĪÉÂļšßĪ·ĪĘĪŦĪÃĪŋĄĨĪģĪÎĪčĪĶĪĘ·ųĩĪÅŠīÄķĪËĪŠĪĪĪÆĪÏĄĪÎēē―ŋåÁĮĪĘĪÉĪÎīÔļĩÅŠĘŠžÁĪÏËÉŲĪËÂļšßĪđĪëĪŽĄĪĪ―ĪÎČūĖĖĄĪŧĀē―ÅŠĪĘē―đįĘŠĪΞŦÁģĪĘđįĀŪĪäĀļÂÖ·ÏĪØĪÎĄÖŧĀē―ÅŠĘŠžÁĄŨĪÎķĄĩëĪÏĪÛĪČĪóĪÉīüÂÔ―ÐÍčĪĘĪĪĄĨĀļĘŠĪÏĪßĪĘīÔļĩÅŠĘŠžÁĪōŧĀē―ĪđĪëĪģĪČĪËĪčĪęĀļĪĪÆĪŠĪęĄĪĪģĪÎĪčĪĶĪĘļÅÃÏĩåīÄķĪÏĄÖąÂĄŨĪČĪĘĪëīÔļĩĘŠžÁĪËĪÏŧö·įĪŦĪĘĪĪĪâĪÎĪĮĪĒĪÃĪŋĪíĪĶĄĨĪ·ĪŦĪ·ĄĪĀļĘŠĪŽĨĻĨÍĨëĨŪĄžĪōÆĀĪëĪŋĪáĪËĪÏĄĪĘŠžÁĪōŧĀē―ĪđĪëšÝĪËĀļĪļĪëīÔļĩÎÏĪÎ―čÍýĪŽÉŽÍŨĪČĪĘĪëĄĨŧĀÁĮļÆĩÛĪËĪŠĪĪĪÆĪÏŧĀÁĮĪŽĪ―ĪÎīÔļĩÎÏĪΞÎĪÆČĪČĪĘĪëĄĨ·ųĩĪīÄķĪËī°ÁīĪËÅŽąþĪ·ĪÆĪĪĪëÎēŧĀīÔļĩķÝĪĮĪĩĪĻĄĪīÔļĩÎÏĪÎ―čÍýĪÎĪŋĪáĪËĄÖÎēŧĀĄŨĪČĪĪĪĶŧĀē―ÅŠĪĘĘŠžÁĪōÉŽÍŨĪČĪđĪëĄĨŧĀÁĮĪŽĪÛĪČĪóĪÉÂļšßĪ·ĪĘĪĪļÅÃÏĩåĪÎĪčĪĶĪĘīÄķĪĮĪÏĄĪīÔļĩÎÏĪōžÎĪÆĪëĪŋĪáĪÎĄÖŧĀē―ÅŠĘŠžÁĄŨĪŽ·įËģĪđĪëĪÎĪĮĪĒĪëĄĨīÔļĩÎÏĪΞÎĪÆČĪōËÉŲĪĘÆóŧĀē―ÚŧĀĪËĩáĪáĪŋĨáĨŋĨóĀļĀŪķÝ°Ęģ°ĪÎĀļĘŠĪÏĄĪĄÖīÔļĩÎÏĪΞÎĪÆČĪηįËģĄŨĪČĪĪĪĶĖäÂęĪËÄūĖĖĪđĪëĪģĪČĪČĪĘĪëĄĨĪÞĪŋĄĪīÔļĩÎÏĪōīðžÁĪÎŧĀē―ÃæīÖĘŠžÁĪËËäĪáĖáĪđĄÖČŊđÚĄŨĪČĪĪĪĶĨĻĨÍĨëĨŪĄžĀļŧš·ÏĪĮĪÏĄĪĪģĪÎĪčĪĶĪĘĄÖīÔļĩÎÏĪΞÎĪÆČĄŨĪÏÉŽÍŨĪĮĪÏĪĘĪĪĪŽĄĪĪ―ĪÎČŊđÚĪōŧŲĪĻĪëĪĀĪąĪÎīðžÁĪÎķĄĩëĪâÅÅŧŌÅÁÃĢ·ÏĪČÚŧĀļĮÄęĮ―ĪōŧýĪÃĪŋÆČÎĐąÉÍÜĀļĘŠĄĪĪÎÂļšßČīĪĪËĪÏđÍĪĻĪéĪėĪĘĪĪĄĨ
ČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏĄĪĘŠžÁĪÎŧĀē―ĪËĪčĪęÆĀĪéĪėĪŋÅÅŧŌĪōļũĨĻĨÍĨëĨŪĄžĪōŧČĪÃĪÆđâĨĻĨÍĨëĨŪĄžē―Ī·ĄĪĪčĪęīÔļĩÅŠĪĘĘŠžÁĪËžõĪąÅÏĪđĪģĪČĪŽ―ÐÍčĪëĪÎĪĮĄĪĪģĪÎĄÖīÔļĩÎÏĪΞÎĪÆČĪηįËģĖäÂęĄŨĪŦĪéĪÏēōĘüĪĩĪėĪÆĪĪĪëĄĨĪ―ĪÎūåĄĪļũĨĻĨÍĨëĨŪĄžĪĮÎēē―ŋåÁĮĪäÎēēŦĪĘĪÉĪōŧĀē―Ī·ĪÆĀļĪĪëĪÎĪĮĄĪÎēŧĀĪĘĪÉĪÎŧĀē―ÅŠÎēēŦē―đįĘŠĪōÂŋÎĖĪËĀļĀŪĪđĪëĪģĪČĪËĪĘĪëĄĨĮÓ―ÐĪĩĪėĪŋÎēŧĀĪÏÎēŧĀīÔļĩķÝĪÎĄÖīÔļĩÎÏĪΞÎĪÆČĄŨĪČĪ·ĪÆÍŅĪĪĪéĪėĪÆĪĪĪŋĪÎĪĀĪíĪĶĄĨĪÞĪŋĄĪČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏīÔļĩÅīĪōļũđįĀŪīðžÁĪČĪ·ĪÆĀļĪĪëĪģĪČĪâēÄĮ―ĪĮĪĒĪëĄĨĪđĪĘĪïĪÁĖĩŧĀÁĮūō·ïēžĪĮÅīĪÎŧĀē―ĪōđÔĪĶĮ―ÎÏĪŽĪĒĪëĪČļĀĪĶĪģĪČĪĮĪĒĪë[21, 22]ĄĨĮÓ―ÐĪĩĪėĪŋŧĀē―ÅīĪÏĄĪĪ―ĪėĪōĄÖīÔļĩÎÏĪΞÎĪÆČĄŨĪČĪ·ĪÆŧČÍŅĪđĪëĀļĘŠĄĘÅīīÔļĩķÝĄËĪËĪČĪÃĪÆĖōĪĀĪÃĪŋĪÐĪŦĪęĪĮĪĘĪŊĄĪģĪÃæĪËÂŋÎĖĪÎÅīĪōÄĀÅÂĪĩĪŧĪëĪģĪČĪËĪĘĪÃĪŋĪÏĪšĪĮĪĒĪëĄĨĪģĪėĪŽĨ·ĨĒĨÎĨÐĨŊĨÆĨęĨĒ―Ðļ―°ĘÁ°ĄĘĪÄĪÞĪęŧĀÁĮČŊĀļĪŽŧÏĪÞĪëÁ°ĄËĪŦĪéžĘūõÅīđÛūēĪČļÆĪÐĪėĪëËÄÂįĪĘŧĀē―ÅīĪÎđÛūēĪŽ·ÁĀŪĪĩĪėĪÆĪĪĪŋļķ°øĪĮĪĒĪíĪĶĄĨ
ĪģĪÎĪčĪĶĪËČóŧĀÁĮČŊĀļ·ŋļũđįĀŪšŲķÝĪÏĄÖŧĀē―ÅŠĘŠžÁĄŨĪŽ·įËģĪ·ĪÆĪĪĪŋļÅÃÏĩåīÄķĪËĪŠĪĪĪÆĄĪĖĩŧĀÁĮūō·ïēžĪĮĪÎļũĪËĪčĪëĘŠžÁŧĀē―ĪōđÔĪĪĄĪÅöŧþĪÎČųĀļĘŠĀļÂÖ·ÏĪËĄÖīÔļĩÎÏĪΞÎĪÆČĄŨĪōÄóķĄĪđĪëĪČĪĪĪĶ―ÅÍŨĪĘĖōģäĪōąéĪļĪÆĪĪĪŋĪÎĪĮĪĒĪíĪĶĄĨĩÕĪËļĀĪĻĪÐĄĪĪâĪ·ôĢĪŦĀÎĪËĀģÂĐĪ·ĪŋĀļĖŋÂÎĄáĀļĘŠĪÎÂÎÆâĪĮĄĪĄÖČóŧĀÁĮČŊĀļ·ŋļũđįĀŪĄŨĪČĪĪĪĶļũĨĻĨÍĨëĨŪĄžĪōÍøÍŅĪ·ĪŋĖĩŧĀÁĮūō·ïēžĪĮĪÎĘŠžÁŧĀē―Ĩ·ĨđĨÆĨāĪÎģŦČŊĪŽĪĘĪŦĪÃĪŋĪČĪ·ĪŋĪéĄĪĘŠžÁ―ÛīÄĨ·ĨđĨÆĨāĪÏūåžęĪŊĩĄĮ―ĪŧĪšĄĪļÅÃÏĩåĪÎČųĀļĘŠĀļÂÖ·ÏĪÏČóūïĪËūŪĪĩĪĘĪâĪÎĪČĪĘĪÃĪÆĪĪĪŋĪĀĪíĪĶĪ·ĄĪļ―šßĪËŧęĪëĀļĖŋŋĘē―ĪÎÆŧÄøĪÏÂÚĪÃĪÆĪ·ĪÞĪÃĪŋĪÏĪšĪĮĪĒĪëĄĨ
ĢļĄĨĪāĪđĪÓĄ§ÍÍĄđĪĘŧëÅĀĪĮĪÎļũđįĀŪŋĘē―ēūĀâ
ĄĄÃÏĩåĀļÂÖ·ÏĪÎČŊÅļĪČĀļĘŠŋĘē―ĪËļũđįĀŪĪŽēĖĪŋĪ·ĪŋĖōģäĄĪĪ―Ī·ĪÆÍŋĪĻĪŋąÆķÁĪÎÂįĪĪĩĪÏ·ŨĪęÃÎĪėĪĘĪĪĪâĪÎĪŽĪĒĪëĄĨÂŋĪŊĪÎļĶĩæžÔĪŽĪģĪÎķËĪáĪÆ―ÅÍŨĪĘĩĄđ―ĪÎĩŊļķĪäŋĘē―ĪËķ―ĖĢĪōžĻĪ·ĪÆĪŠĪęĄĪÍÍĄđĪĘŧëÅĀĪŦĪéļũđįĀŪĩĄĮ―ĪÎŋĘē―ēáÄøĪŽÏĀĪļĪéĪėĪÆĪĪĪëĄĨĪ―ĪÎĪčĪĶĪĘŋĘē―ēūĀâĪÎÃæĪŦĪéĄĪĪĪĪŊĪÄĪŦĪÎķ―ĖĢŋžĪĪĘļļĨĪōĨÔĨÃĨŊĨĒĨÃĨŨĪ·ĄĪĘļļĨĨęĨđĨČĪΚĮļåĪËēÃĪĻĪÆĪŠĪĪĪŋĄĨËÜĀÛÏĀĪōÄŊĪáĪÆĄĪĖĪĪĀÁīÍÆĪŽĖĀĪŦĪČĪĘĪÃĪÆĪĪĪĘĪĪĄÖļũđįĀŪĪÎŋĘē―ĄŨĪËīØĪ·ĪÆĄĪÂŋūŊĪĘĪęĪČĪâķ―ĖĢĪōŧýĪŋĪėĪŋÆÉžÔĪÏĀ§ČóĪČĪâĪģĪėĪéÍĨĪėĪŋĘļļĨĪōžęĪËžčĪÃĪÆĪīÍũÄšĪĪŋĪĪ[23-27]ĄĨ°úÍŅĘļļĨ
1. Margulis, L. Symbiotic Planet:: Basic Books Publishers, New York, 1998.2. Garrity, G. M. and Holt, J. G. The road map to the Manual: p. 119-155 In D. R. Boone, R. W. Castenholz and G. M. Garrity, (ed.), Bergey's Manual of Systematic Bacteriology, 2nd edn. vol. 1: Springer-Verlag, New York, 2001.
3. .Hanada, S. and Pierson, B. K. The Family Chloroflexaceae, In M. Dworkin, K. H. Schleifer and E. Stackebrandt, (ed.), The Prokaryotes: An Evolving Electronic Resource for the Microbiological Community, 3rd edition (release 3.11, November 22 2002): Springer-Verlag, New York, 2002.
4. Pierson, B. K. and Castenholz, R. W. Taxonomy and physiology of filamentous anoxygenic phototrophs, p. 31-47 InR. E. Blankenship, M. T. Madigan and C. E. Bauer, (ed.): Anoxygenic Photosynthetic Bacteria. Kluwer Academic Publishers, Dordrecht, 1995.
5. Imhoff, J. F. Taxonomy and physiology of phototrophic purple bacteria and green sulfur bacteria, p. 1-15 InR. E. Blankenship, M. T. Madigan and C. E. Bauer, (ed.), Anoxygenic Photosynthetic Bacteria: Kluwer Academic Publishers, Dordrecht, 1995.
6. Imhoff, J. F. The anoxygenic photorrophic purple bacteria, p. 631-637 InD. R. Boone, R. W. Castenholz and G. M. Garrity, (ed.), Bergey's Manual of Systematic Bacteriology, 2nd edn. vol. 1: Springer-Verlag, New York, 2001.
7. 0vermann, J. Family I. "Chlorobiaceae," Green Sulfur Bacteria, p. 601-623 InD. R. Boone, R. W. Castenholz and G. M. Garrity, (ed.), Bergey's Manual of Systematic Bacteriology, 2nd edn. vol. 1: Springer-Verlag, New York, 2001.
8. Madigan, M. T. and Ormerod, J. G. Taxonomy, physiology and ecology of Heliobacteria, p. 17-30 InR. E. Blankenship, M. T. Madigan and C. E. Bauer, (ed.), Anoxygenic Photosynthetic Bacteria: Kluwer Academic Publishers, Dordrecht, 1995.
9. Madigan, M. T. Family VI. "Heliobacteriaceae" Beer-Romero and Gest 1987,113, p. 625-630 InD. R. Boone, R. W. Castenholz and G. M. Garrity, (ed.), Bergey's Manual of Systematic Bacteriology, 2nd edn. vol. 1: Springer-Verlag, New York, 2001.
10. Clayton, R. K. Photosynthesis, Physical mechanisms and chemical patterns: Cambridge University Press, Cambridge, 1980.
11. Blankenship, R. E. and Prince, R. C. Excited-state redox potentials and Z scheme of photosynthesis, Trends Biochem. Sci. 10, 382-383 (1985).
12. Olson, J. M. and Pierson, B. K. Evolution of reaction centers in photosynthetic prokaryotes, International Review of Cytology 108, 209-248 (1987).
13. Dovindjee and Coleman, W. J. How plants make oxygen, Scientific American 262, 50-58 (1990).
14. Michel, H. and Deisenhofer, J. Relevance of the photosynthetic reaction center from purple bacteria to the structure of photosystem II, Biochemistry 27, 1-7 (1988). 15. Pierson, B. K. and Thornber, J. P. Isolation and spectral characterization of
photochemical reaction centers from the thermophilic green bacterium Chloroflexus aurantiacus strain J-10-fl, Proc. Natl. Acad. Sci. USA 80, 80-84 (1983).
16. Krauss, N., Hinrichs, W., Witt, I., Fromme, P., Pritzkow, W., Dauter, Z., Betzel, C., Wilson, K. S., Witt, H. T. and Saenger, W. 3-dimensional structure of system I of photosynthesis at 6 angstrom resolution, Nature 361, 326-331 (1993).
17. Xiong, J., Fischer, W. M., Inoue, K., Nakahara, M. and Bauer, C. E. Molecular evidence for the early evolution of photosynthesis, Science 289, 724-1730 (2000).
18. Garcia-Gil, L. J., Gich, F. B. and Fuentes-Garcia, X. A comparative study of bchG from green photosynthetic bacteria, Arch. Microbiol. 179, 108-115 (2003).
19. Raymond, J., Zhaxybayeva, O., Gogarten, J. P., Gerdes, S. Y. and Blankenship, R. E. Whole-genome analysis of photosynthetic prokaryotes, Science 298, 1616-1620 (2002).
20. Holland, H. D. Early Proterozoic atmospheric change, p. 237-244 InS. Bengston, (ed.), Early life on Earth: Columbia University Press, New York, 1994.
21. Ehrenreich, A. and Widdel, F. Anaerobic oxidation of ferrous iron by purple bacteria, a new-type of phototrophic metabolism, Appl. Environ. Microbiol. 60, 4517-4526 (1994).
22. Heising, S., Richter, L., Ludwig, W. and Schink, B. Chlorobium ferrooxidans sp nov., a phototrophic green sulfur bacterium that oxidizes ferrous iron in coculture with a "Geospirillum" sp strain, Arch. Microbiol. 172, 116-124 (1999).
23. Blankenship, R. E. Origin and early evolution of photosynthesis, Photosynth. Res. 33, 91-111 (1992).
24. Blankenship, R. E. The origin and evolution of oxygenic photosynthesis, TIBS 23, 94-97 (1998).
25. Lockhart, P. J., Steel, M. A. and Larkum, A. W. D. Gene duplication and the evolution of photosynthetic reaction center proteins, FEBS Letters 385, 193-196 (1996).
26. Xiong, J. and Bauer, C. E. Complex evolution of photosynthesis, Annual Review of Plant Biology 53, 503-521 (2002).
27. ūūąšđîČþ, °ËÆĢ ČË ŧĀÁĮÂįĩĪĪČĀļĖŋĪÎĀąĪōĪâĪŋĪéĪ·ĪŋļũđįĀŪĄ―Ī―ĪÎÃÂĀļĪČŋĘē―ĪÎĨ·ĨĘĨęĨŠ, ēĘģØ 68, 839-844 (1998).
28. Hasegawa, M. and Kishino, H. Accuracies of the simple methods for estimating the bootstrap probability of a maximumlikelihood tree, Mol Biol Evol 11, 142-145 (1994).